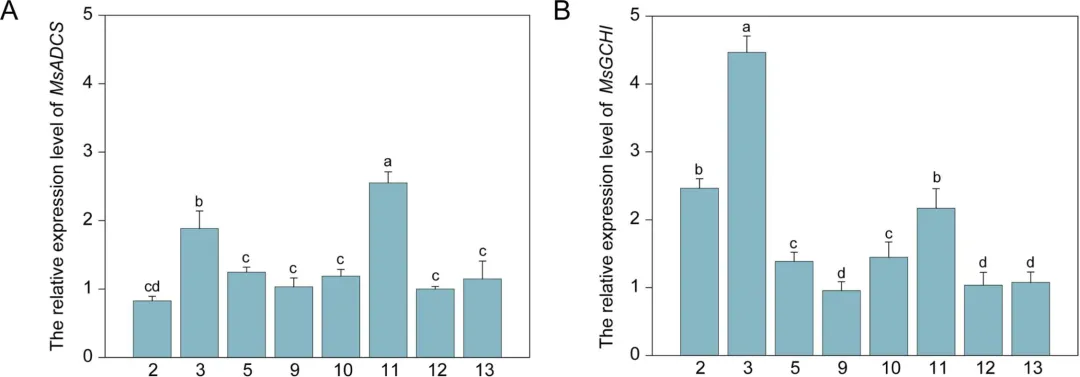
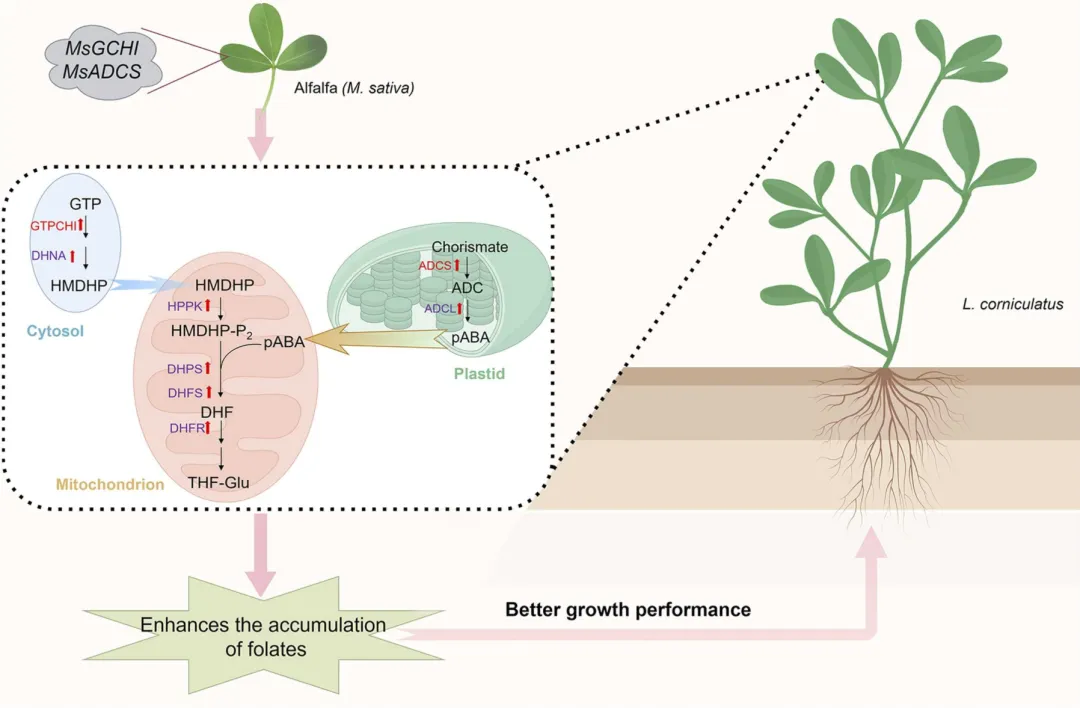
研究人员首先通过农杆菌介导法将来自紫花苜蓿的叶酸合成关键酶基因MsGCHI和MsADCS转入百脉根,经Basta筛选和PCR鉴定获得13个阳性转基因株系。利用RT-PCR和RT-qPCR检测后,最终筛选出5个双基因高表达株系(GA-2、3、5、10、11)用于后续试验(图1)。
图1 MsADCS (A)和MsGCHI (B)在转基因百脉根中的表达分析
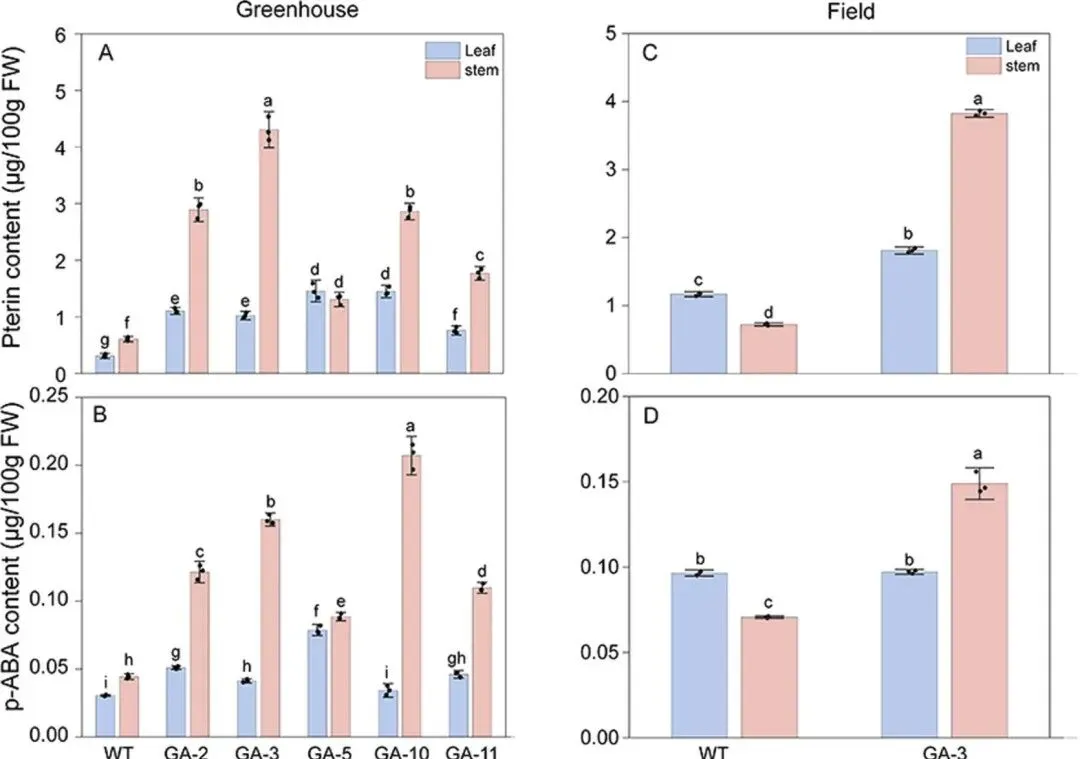
为评估MsGCHI和MsADCS共表达对转基因百脉根叶酸代谢的影响,研究人员利用HPLC-MS/MS分别检测了温室和大田条件下转基因株系叶片和茎中两种叶酸前体(喋呤和对氨基苯甲酸(p-ABA))的含量。结果显示,转基因株系叶片和茎中喋呤含量较对照分别提高2.7倍和3.3倍,p-ABA含量分别增加1.1倍和4.7倍(图2A-B)。田间条件下,GA-3株系叶片和茎中喋呤含量较野生型分别提高0.4倍和4.3倍,p-ABA含量变化趋势与温室条件一致(图2C-D),证实了环境波动下代谢改良效果的稳定性。
图2 温室和大田条件下不同株系百脉根中叶酸前体(喋呤和对氨基苯甲酸(p-ABA))含量
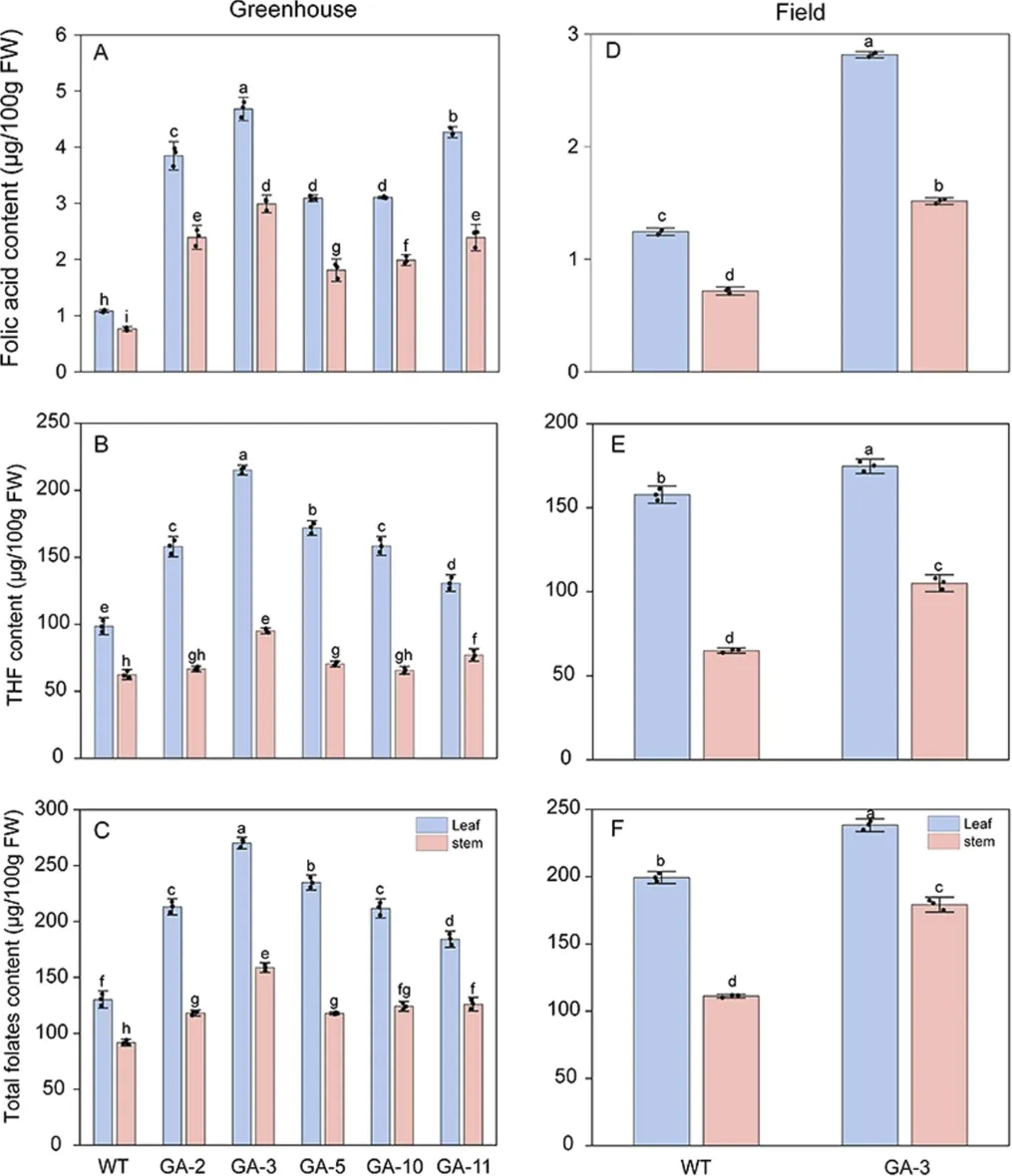
研究进一步对温室和大田条件下的叶酸衍生物及总叶酸含量进行了系统分析。温室条件下,转基因株系叶片和茎中多种叶酸衍生物显著积累:叶酸含量(氧化产物)较对照平均提高2.5倍和2.0倍(图3A);THF相关衍生物作为主要组分,在GA-3株系分别提高1.2倍和0.5倍(图3B);此外,5-M-THF和5-F-THF含量显著增加,GA-5株系叶片中5-M-THF提高2.5倍,GA-3株系叶片和茎中5-F-THF分别提高1.4倍和1.7倍。总叶酸含量在转基因株系叶片和茎中较野生型分别提高1.4-2.1倍和1.3-1.7倍(图3C)。田间条件下,GA-3株系叶片和茎中总叶酸含量较野生型分别提高1.2倍和1.6倍,各衍生物积累模式与温室基本一致(图3D-F)。
图3 温室和大田条件下不同株系百脉根中叶酸、四氢叶酸相关衍生物和总叶酸含量
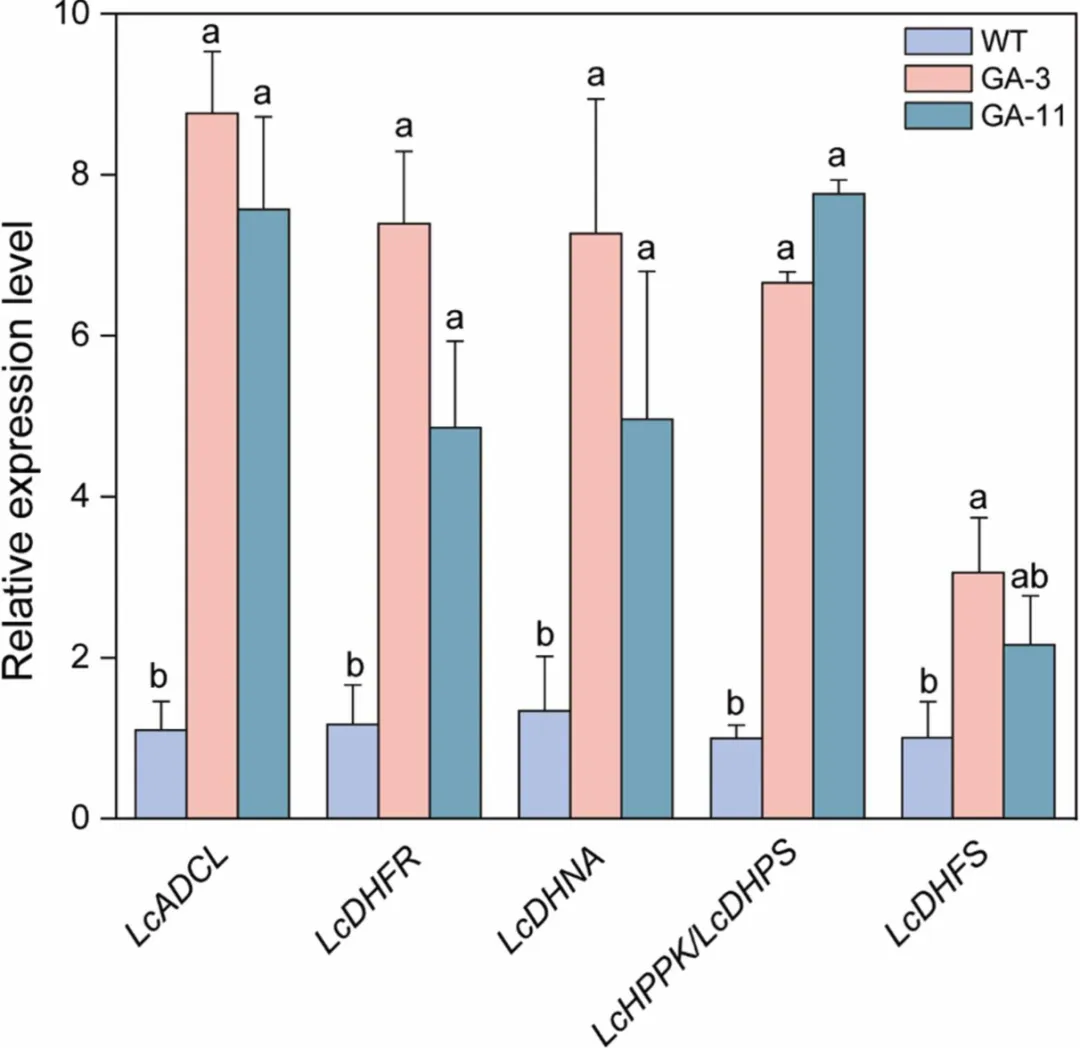
为探究MsGCHI和MsADCS共表达对内源叶酸合成途径的转录调控,研究人员对GA-3和GA-11株系中6个关键酶基因(ADCL、HPPK/DHPS、DHFR、DHFS、DHNA)进行RT-qPCR分析。结果显示,GA-3株系所有基因表达均显著上调;GA-11株系中5个基因显著上调,仅DHFS变化不显著(图4),表明外源基因导入激活了内源叶酸合成通路的协同响应。
图4 温室条件下野生型和两个转基因百脉根株系叶片中叶酸生物合成途径关键酶基因的RT-qPCR分析
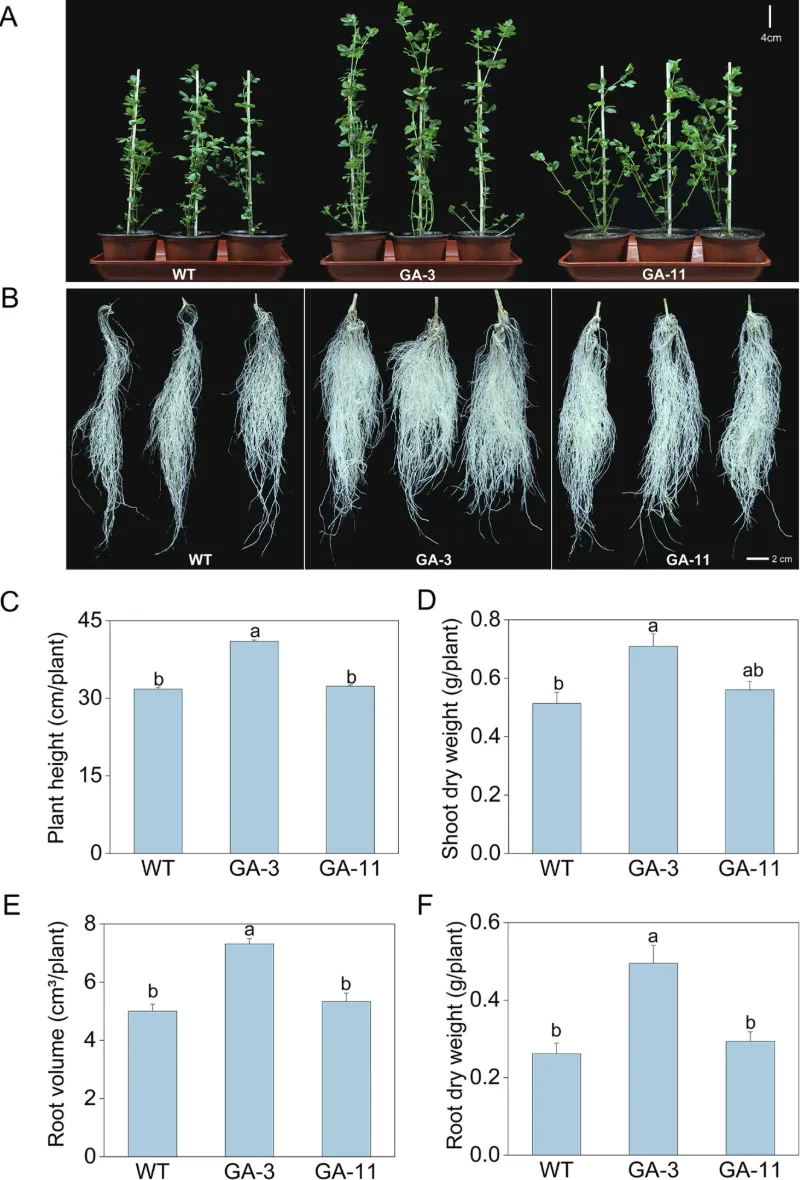
为评估转基因植株的生长表现,研究人员对温室和大田条件下的转基因株系进行表型分析。温室条件下,4周龄转基因株系长势优于野生型(图5A),其中GA-3株系株高、地上部干重、根体积和根干重分别较野生型提高29.2%、38.2%、34.9%和89.2%(图5B-F)。
图5 温室条件下野生型和转基因百脉根株系的生长表型分析
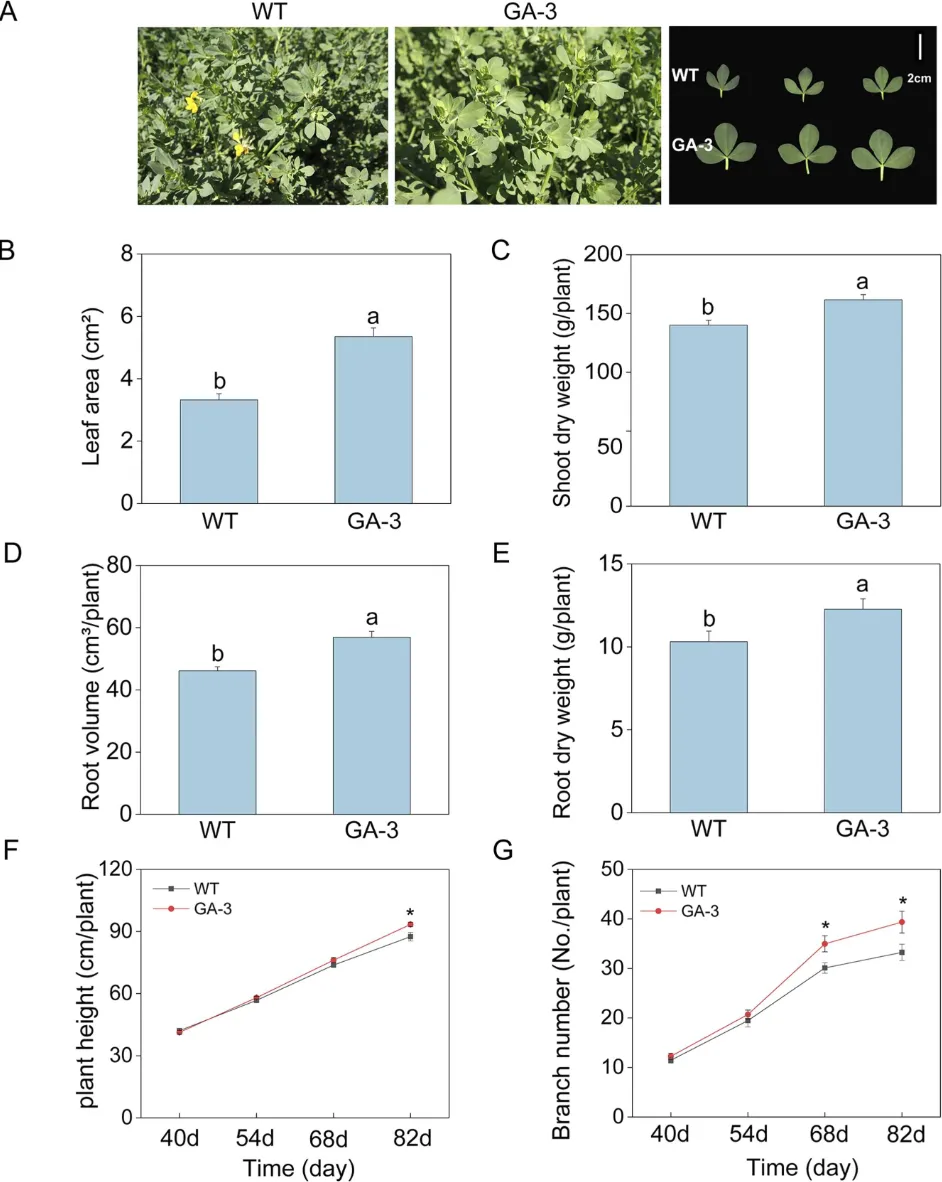
田间条件下,转基因植株维持生长优势并出现形态变异:GA-3叶片呈倒卵形、先端圆钝,野生型为斜卵形、先端尖(图6A);初花期GA-3叶面积、地上部干重、根体积和根干重较野生型分别显著增加61.2%、15.3%、23.3%和18.9%(图6B-E);移栽82天后GA-3株高和分枝数均高于野生型(图6F-G)
图6 大田移栽后野生型和转基因百脉根株系的生长表型分析





 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
