Food Chem. 中国农业科学院-食品中玉米赤霉烯酮降解用高耐热耐酸突变型玉米赤霉烯酮内酯水解酶的合理设计
2025 年 11 月 9 日,来自中国农业科学院的Lanxue Li和Yingguo Bai等人在Food Chemistry上发表了一篇题为Rational design of an engineered zearalenone lactone hydrolase with enhanced thermostability and acid tolerance for zearalenone degradation in food的研究性论文。
Abstract
玉米赤霉烯酮(ZEN)是一种在谷物中常见且稳定的镰刀菌霉菌毒素,对食品安全构成威胁,且常规加工方法难以将其去除。天然的ZEN水解酶在工业加热和酸性条件下往往效果不佳。本研究通过一种混合计算策略,整合了能量计算、静电势优化、共有序列分析和二硫键设计,成功改造出一个稳健的变体R76Q/T217C-G242C。所得的生物催化剂能在30分钟内完全降解16.6 mg/L的ZEN,在55°C下处理5分钟后仍保留71%的活性(提升了10倍),在pH 5.0条件下活性超过75%,且在40°C下的半衰期为192.5分钟。该变体还能去除霉变酒糟中95.13%的天然ZEN。结构和动力学分析揭示了其通过设计的二硫键和优化的相互作用网络介导的稳定机制。这项研究为在挑战性的食品加工环境中降解ZEN提供了一种高性能的酶学解决方案。
Introduction
真菌毒素是由产毒真菌产生的有毒次级代谢产物,持续威胁着全球食品安全、农业生产和牲畜健康。在已发现的400多种真菌毒素中,玉米赤霉烯酮(ZEN;C₁₈H₂₂O₅)因其强效的内分泌干扰效应及在食品加工过程中的稳定性,已被欧洲食品安全局列为优先控制污染物。ZEN广泛污染全球谷物,据报道埃及玉米中的检出浓度高达38,400微克/千克,南斯拉夫和南非的样品中甚至超过10,000微克/千克,凸显其普遍存在性及有效治理的迫切需求。
从结构上看,ZEN的大环δ-内酯和共轭双键体系与17β-雌二醇高度相似,能够高亲和力结合雌激素受体并激活雌激素活性。这种分子模拟效应使其即使在低暴露水平下也能产生多器官毒性。在最为敏感的幼年猪模型中,饲料中仅含1-5毫克/千克的ZEN即可诱发外阴肿胀、子宫增大和不孕症。暴露于10-30毫克/千克ZEN的啮齿类动物则表现出精子发生受损、孕酮水平降低和产仔数减少。人类流行病学研究进一步将ZEN暴露与波多黎各及匈牙利儿童的性早熟相关联,凸显其公共健康风险。此外,ZEN还具有遗传毒性和潜在致癌性,加剧了其在食物链中的风险。
传统的物理和化学脱毒方法常因ZEN固有的稳定性而难以彻底去除,且存在能耗高、营养素损失或产生有毒副产物等缺陷。因此,开发既能选择性破坏ZEN毒性骨架又能确保降解产物生物安全性的靶向降解技术,对提升食品安全水平至关重要。
基于微生物源特异性内酯水解酶的酶促生物脱毒技术,因其高特异性、温和反应条件和环境兼容性,已成为前景广阔的替代方案。这些酶能够催化ZEN内酯环的裂解,生成经结构确认无雌激素活性的开环产物。然而,天然ZEN水解酶的实际应用因热稳定性和耐酸性不足而严重受限——这与其在食品加工中常见的高温及酸性环境不相适应。
为突破这一瓶颈,提出假设:通过整合基于能量、静电作用、进化信息和结构强化原则的多维计算设计策略,可在不牺牲催化效率的前提下协同提升酶的热稳定性和耐酸性。既往的酶工程改造多聚焦于单一稳定性参数,常导致稳定性-活性权衡效应,而本研究的整合策略能够实现稳健酶变体的系统性设计。应用该技术体系,成功构建了R76Q/T217C-G242C突变体,该变体展现出显著增强的热稳定性和酸稳定性,为工业食品脱毒提供了高性能生物催化剂。这项研究不仅破解了关键稳定性瓶颈,更为开发下一代真菌毒素降解酶建立了普适性工程范式。
结果
在食品安全领域,谷物中的真菌毒素污染,特别是玉米赤霉烯酮(ZEN)污染,对公众健康和食品质量构成重大威胁。前期研究从美洲大蠊中鉴定出一种新型ZEN内酯水解酶ZHD607(GenBank登录号:KIW70607),该酶具有优良的脱毒活性。通过理性设计,后续引入酪氨酸残基构建了I160Y突变体,以增强与ZEN大环内酯环的疏水作用和π-π堆积相互作用。该突变体比活力达16,652 U/mg,可在30分钟内完全降解16.6 mg/L的ZEN,催化效率较ZHD607提升3.4倍。然而,其适中的热稳定性限制了工业应用——在40℃下半衰期仅70.7分钟。为攻克这一瓶颈,启动了系统性工程改造研究,旨在开发适用于工业化食品加工的稳健酶变体。
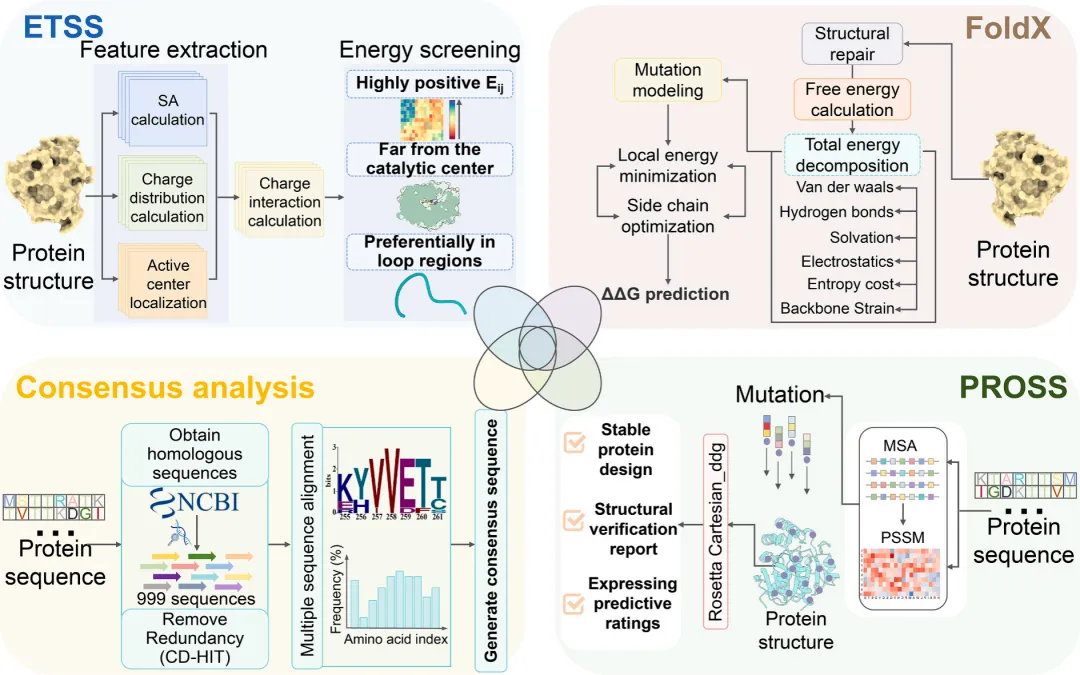
1. 玉米赤霉烯酮内酯水解酶耐热突变体的计算机辅助设计
建立了一个四维正交设计框架,用于对I160Y母体进行稳定性改造(图1),旨在平衡进化适应性、热力学稳定性和结构兼容性(图1)。该整合策略包括:(1)通过ETSS进行静电优化,识别出34个热不稳定残基;(2)对999个同源序列进行进化保守性分析,筛选出37个保守位点;(3)利用FoldX进行全局稳定性扫描,预测出176个稳定化突变(ΔΔG < 0 kcal/mol);(4)通过PROSS进行进化可塑性图谱分析,锁定50个功能兼容的替代位点。
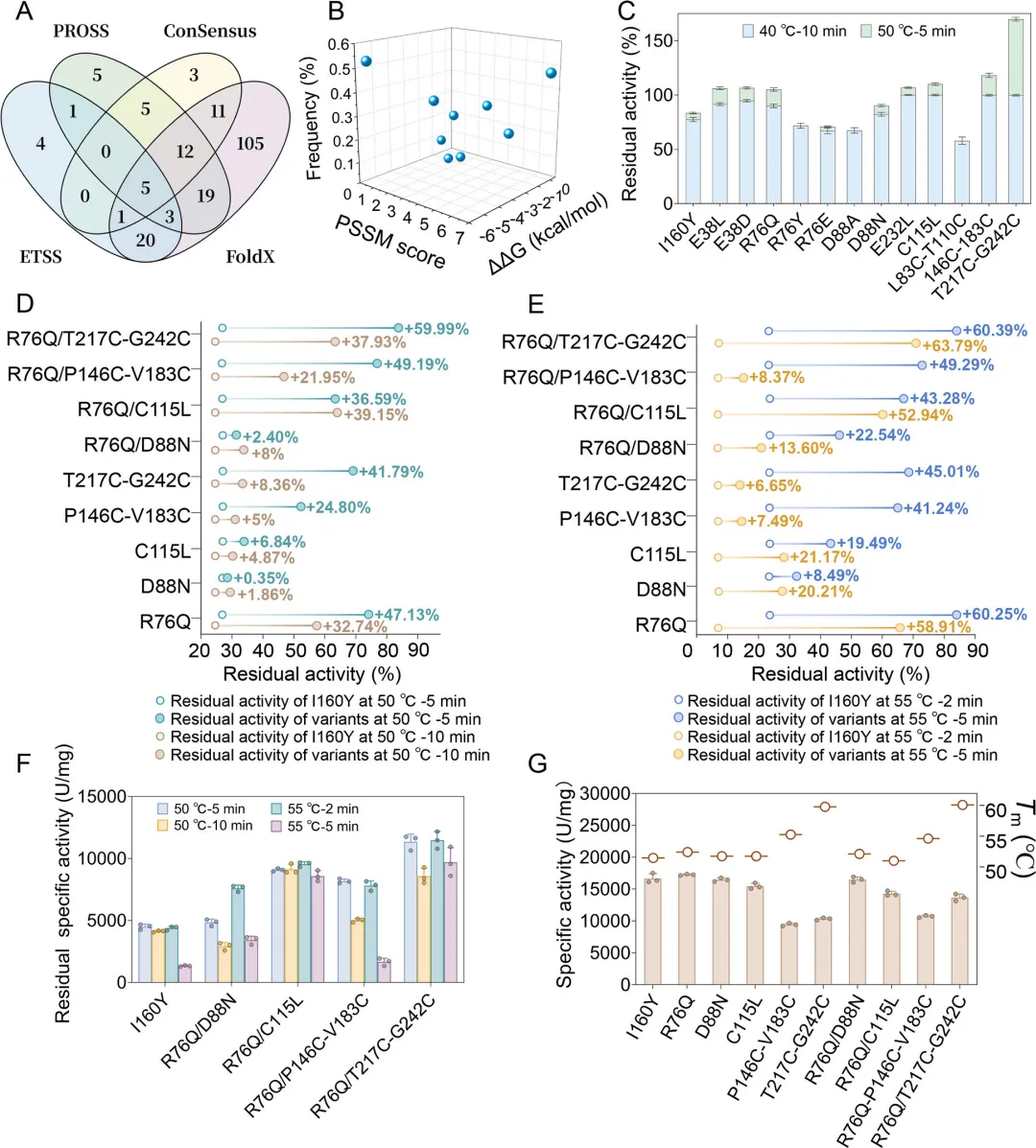
为克服ETSS模型在确定最优突变类型方面的局限性,采用了以位点为中心的筛选策略。通过多维保守性分析对候选突变进行掩码处理(例如用"M1"替代具体氨基酸突变),以优先考虑四个模块间的位点重叠区域。维恩图整合分析确定了五个高置信度的失稳热点(E38、R76、D88、C115和E232)(图2A)。随后结合热力学稳定性评估与系统发育保守性验证,获得了九个生化性质优良的突变体:E38D/L、R76Q/E/Y、D88N/A、C115L和E232L(图2B)。
同时,运用互补的计算工具进行了二硫键工程改造。Discovery Studio 2017软件筛选出22个几何构型可行的候选位点,而Disulfide by Design 2.0则根据热力学阈值筛选出7个符合条件的突变。通过交叉分析,获得了同时满足空间构象和能量要求的三个共有二硫键突变体(P146C-V183C、T217C-G242C和L83C-T110C)。这三个二硫键突变体与四维正交框架获得的9个单点突变体,共同构成了包含12个候选突变体的最终实验验证库。
2. 候选突变体的表达与热稳定性表征
首先采用粗酶提取液在40℃条件下处理10分钟,对12个候选突变体的热稳定性进行初步筛选。结果显示,9个突变体的残余酶活维持在50%以上,而R76Y、R76E、D88A和L83C-T110C突变体的稳定性较I160Y母体酶有所降低(图2C,p < 0.05)。对表现优异的突变体进行纯化以开展详细表征,而E38D、E38L和E232L因表达产量低被排除。
通过50℃和55℃条件下的延长时间热处理实验,筛选出五个优势突变体:R76Q、D88N、C115L、P146C-V183C和T217C-G242C(图2D、E)。其中,R76Q突变体表现出最高的热稳定性,在55℃处理2分钟和5分钟后分别保持84%和66%的残余活性,较I160Y分别提升2.5倍和8.4倍。含二硫键的突变体P146C-V183C和T217C-G242C在50℃处理5分钟后残余活性分别为65%和69%,较I160Y分别提高92%和155%。虽然R76Q、D88N和C115L保持了较高的比活力,但含二硫键的突变体比活力出现适度下降(约为I160Y的62.5%),却获得了构象稳定性提升,其熔解温度(ΔTm)分别增加3.78℃和8.28℃(图2G)。这表明二硫键工程在保留关键催化功能的同时,优先实现了结构完整性的强化。
3. 高耐热突变体的组合突变构建
鉴于R76Q突变体优异的热稳定性及其在食品工业中的应用潜力,以其为母本构建了组合突变体(R76Q/D88N、R76Q/C115L、R76Q/P146C-V183C、R76Q/T217C-G242C)。与单一突变体相比,所有双突变体在热稳定性和催化活性方面均表现出协同增强效应(图2D、E)。其中,R76Q/T217C-G242C突变体表现尤为突出:在50℃处理5分钟后,其残余活性较I160Y提升3.1倍(82.94% vs. 26.96%),处理10分钟后仍保持2.5倍优势(62.69% vs. 24.77%);在55℃严苛条件下,处理2分钟时活性提升3.6倍(83.91% vs. 23.53%),处理5分钟后更提升至10.0倍(70.92% vs. 7.13%)。值得注意的是,R76Q/T217C-G242C在保持I160Y母体酶82%催化效率的同时,实现了热稳定性的显著提升。通过残余比活力分析进一步量化其稳定性优势:在50℃处理5分钟和10分钟后,该突变体较I160Y分别提升2.53倍和2.08倍;在55℃处理2分钟和5分钟后,则分别提升2.62倍和7.29倍(图2F,p < 0.05)。
为系统评估R76Q/T217C-G242C突变带来的稳定性增益,对其进行了动力学和热力学稳定性参数表征。40℃条件下的热失活动力学分析显示,R76Q/T217C-G242C突变体的功能半衰期(40℃,pH=9.0时t1/2 = 192.5 ± 5.3分钟)较I160Y(40℃,pH=9.0时t1/2 = 70.7 ± 2.9分钟)延长了121.8分钟。热力学稳定性分析表明,其熔解温度提升8.58 ± 0.23℃(60.12℃ vs. 51.54℃)(图2G,p < 0.05)。这些结果证实了在热胁迫条件下该突变体酶结构具有协同稳定效应。
4. pH稳定性评估
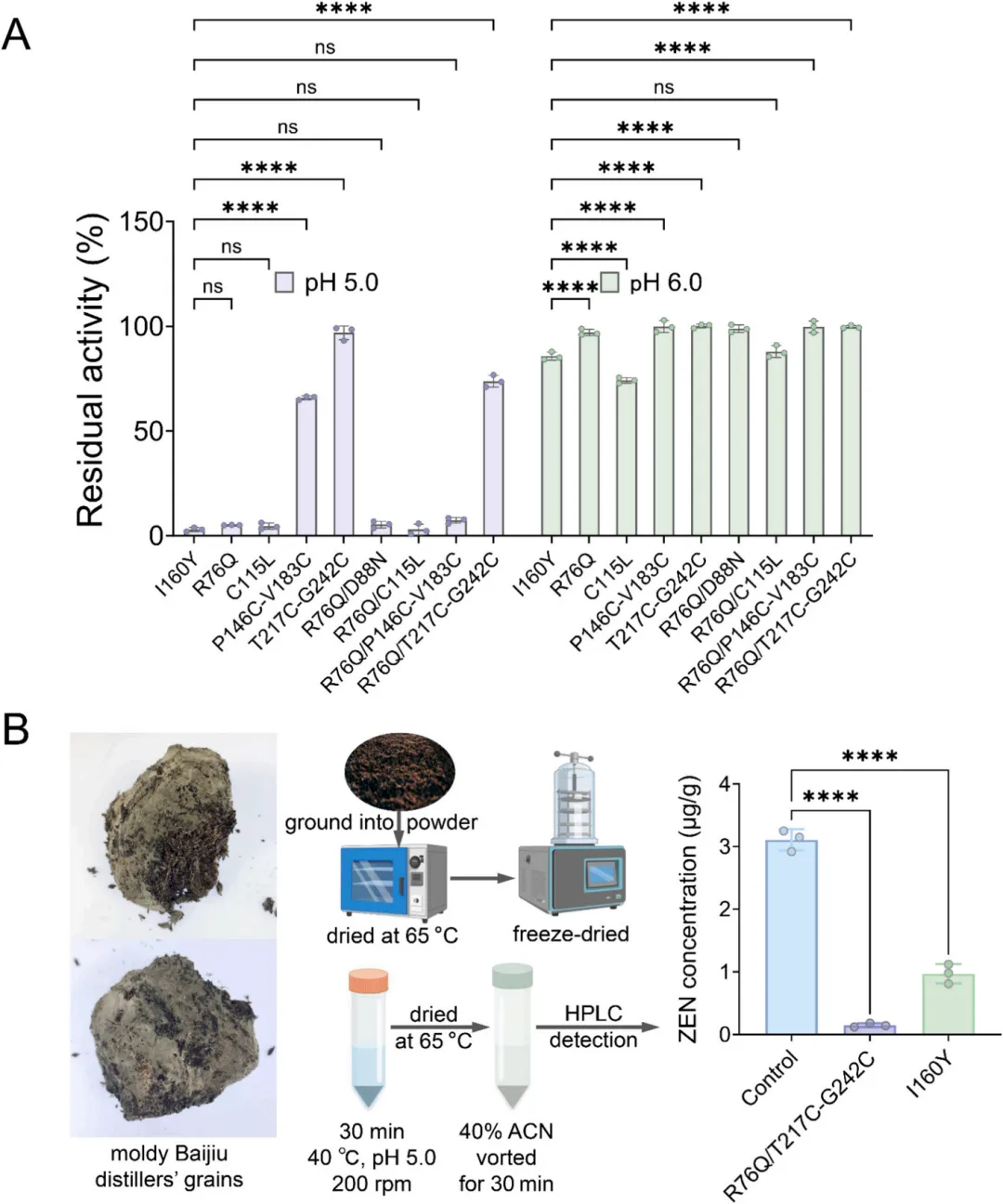
考虑到食品种类的多样性,特别是发酵食品和饮料中的酸性环境,评估了I160Y及其突变体在pH 5.0–6.0条件下的pH稳定性。I160Y在pH 5.0条件下处理1小时后仅保留3%的活性,证实了其酸敏感性。除C115L外,所有突变体的耐酸性均得到改善。其中,含二硫键的突变体T217C-G242C在pH 5.0–6.0范围内保持完全活性,而组合突变体R76Q/T217C-G242C在pH 5.0条件下处理1小时后残余活性超过75%(图3A,p < 0.0001)。
为阐明耐酸性提升的分子机制,利用H++程序在pH 5.0、离子强度0.15 mol/L条件下分析了R76Q/T217C-G242C突变对催化三联体(Ser105-His243-Glu129)pKa值的影响。结果表明,与I160Y相比,R76Q/T217C-G242C中His243的pKa值降低0.3个单位,表明通过调控催化残基的pKa值可增强酶的pH适应性。这一调控机制为理性酶工程改造提供了新策略:通过设计调节催化残基pKa值的突变,可间接提升酶的pH适应性,以满足更广泛的工业应用需求。
5. 优势突变体在霉变白酒酒糟中的应用研究
为评估工程化改造的R76Q/T217C-G242C突变体在真实且具挑战性的食品链场景中的实际应用效果,选取霉变白酒酒糟进行ZEN降解试验。选择该基质主要基于两点原因:(1)酒糟作为白酒生产的主要副产物,因其营养丰富且常在温暖潮湿环境下储存,极易受真菌毒素污染,是高风险饲料原料;(2)该发酵基质固有的酸性特性对多数ZEN降解酶构成严峻挑战,为验证R76Q/T217C-G242C突变体的耐酸性提供了严苛的测试环境。
在与食品加工相关的模拟酸性条件(pH 5.0)下,R76Q/T217C-G242C突变体处理30分钟后,对酒糟中天然存在的ZEN降解率达95.13%,使其浓度从初始的3.08微克/克降至0.15微克/克(图3B)。相比之下,在相同条件下用母体酶I160Y处理的对照组,其降解率显著较低,仅为69.22%(p < 0.0001),ZEN浓度降至0.95微克/克。这意味着经R76Q/T217C-G242C突变体处理后,ZEN终浓度较I160Y组降低6.3倍(0.15微克/克 vs. 0.95微克/克)。这些结果证明该突变体在复杂真实基质中具有更强的降解能力,凸显了其在提升受污染谷物及其副产物安全性方面的工业应用潜力。
6. 增强热稳定性的结构基础
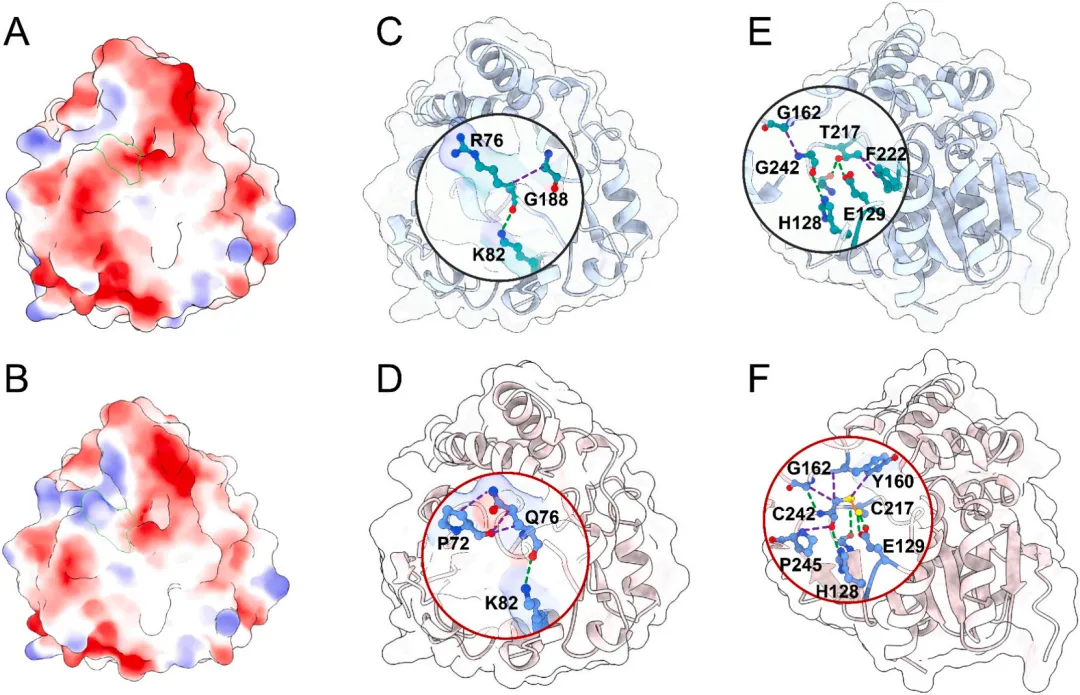
R76Q/T217C-G242C突变体热稳定性的提升源于两个关键突变位点相互作用网络的重构。在76位点,R76Q突变在保留R76 O与K82 NZ之间关键极性接触(2.875 Å,与I160Y中观察到的结构类似)的同时,将原有的R76 CA与G188 CA之间的疏水相互作用(4.115 Å)优化为Q76 NE2与P72 CB之间更有利的疏水接触(3.862 Å)(图4C、D)。此外,V78 CG2与Q76主链N原子之间新形成的疏水相互作用(3.768 Å)进一步稳定了侧链构象。Q76主链羰基氧与D77之间的偶极-偶极相互作用(3.234 Å)通过静电作用限制主链柔性,与邻近疏水接触共同促进环区结构的稳定性。除局部相互作用外,R76Q突变还诱导酶分子静电表面发生有利重构。表面电荷分析显示,I160Y中R76位点周围呈现显著的正静电势,形成潜在排斥性微环境(图4A)。R76Q突变有效缓解了这种电荷过饱和状态,形成更中性和互补的静电表面(图4B),从而增强结构完整性与稳定性。
T217C-G242C二硫键突变通过协同重构疏水网络和氢键网络,显著提升了酶的热稳定性(图4E、F)。C217与C242之间形成了稳定的分子内二硫键(1.9 Å),替代了I160Y中T217与G242之间较弱的极性接触。这一共价连接诱导了邻近疏水相互作用的系统性重塑:C242的硫原子与Y160苯环形成π-π堆积作用(3.930 Å),增强了范德华相互作用;C242 CA与G214 CA之间形成跨β-折叠疏水桥(4.082 Å),同时C242 CB与G162 CA的紧密堆积(4.025 Å)强化了β-折叠与环区的连接。值得注意的是,P245吡咯烷环与C242主链氧原子之间的疏水相互作用(3.505 Å)填充了结构空腔,抑制了环区波动。这些作用共同提升了疏水相互作用的密度和能量增益。氢键网络也得到优化:C217硫原子与E129 OE1(2.877 Å)和OE2(3.102 Å)形成双重氢键,替代了I160Y中单一的T217 OG1-E129 OE1氢键。关键主链氢键(C217 N-G214 O,2.539 Å;C242 O–H128 NE2,3.149 Å)相较于I160Y(分别为3.176 Å和3.266 Å)键长分别缩短19.8%和5.6%,显著增强了结构刚性。
7. 增强热稳定性的动态分子机制
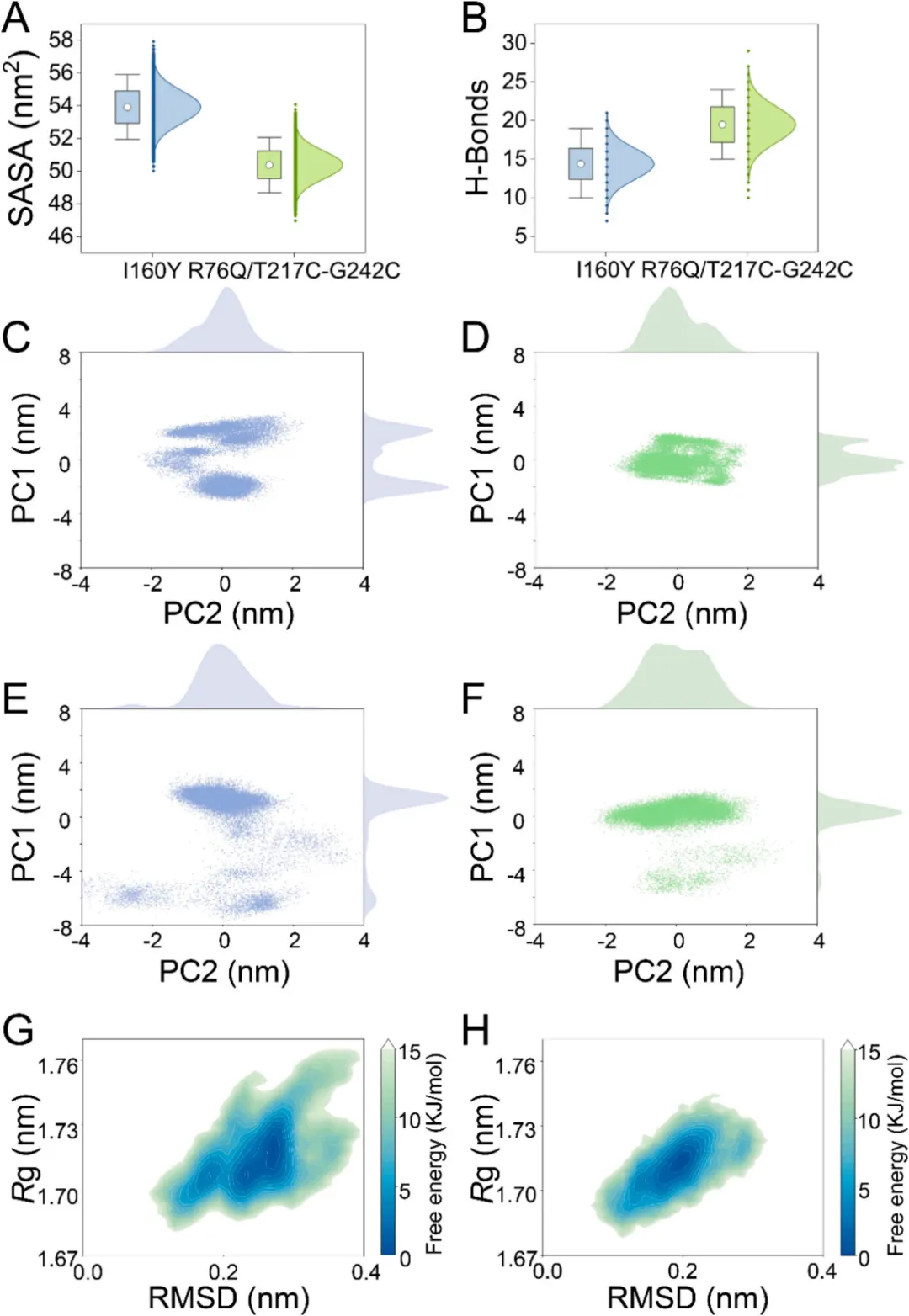
为阐明这些结构修饰的动态效应,进行了300 K条件下200 ns的分子动力学模拟,比较了突变体与I160Y的差异。全局构象分析显示,R76Q/T217C-G242C的RMSD波动(0.2 nm)较I160Y(0.275 nm)减小,表明热稳定性增强。自由能形貌分析进一步证实这一发现:突变体占据单一明确的低能态盆地,回转半径(1.69–1.73 nm)和RMSD(0.20–0.27 nm)范围狭窄,而I160Y则采样多个亚稳态(图5G、H)。局部结构稳定化体现在RMSF图谱中:该图谱识别出I160Y中四个高柔性区域,包括区域2(R66-W86)和区域4(T217-G242)——正是突变体的突变位点所在。这些区域溶剂可及表面积降低和氢键网络增强进一步证实了靶向稳定化效应(图5A、B)。
主成分分析和动态交叉相关矩阵进一步揭示了别构偶联机制。构象聚类分析显示,突变体在区域2和区域4呈现集中运动采样,而I160Y则分散分布于三个聚类簇中(图5C-F)。DCCM分析表明,突变体中76号残基与R5-V20区域之间、242号残基与R5-V20及R55-A70区域之间的正动态相关性显著增强,表明跨结构域相互作用网络得到强化。同时,突变体中较弱的负相关性反映出相较于I160Y,其去稳定化运动有所减少。这些多尺度观察结果——从局部相互作用到全局构象动力学——共同表明,突变体热稳定性的提升源于协同效应:易感结构元件的靶向稳定化、结构域协调性改善以及有害运动的抑制。这种分层稳定机制为食品相关酶的理性工程改造提供了设计蓝图。
Discussion
酶法脱除玉米赤霉烯酮(ZEN)的工业化应用潜力长期受限于天然内酯水解酶在食品加工条件下的固有不稳定性。通过酶工程改造突破这些限制条件构成了关键性进展,对于保障乳制品和谷物制品安全尤为重要。本研究建立了一个多维计算框架,通过协同整合四种正交设计策略,实现了对传统单参数优化的根本性突破,有效规避了稳定性与活性之间的典型权衡效应。该框架融合了基础热力学认知与机器学习辅助预测:例如,FoldX能量计算量化了R76Q取代的稳定化贡献(ΔΔG = −0.779 kcal/mol),该预测与实验数据高度吻合;ETSS分析则诊断出R76残基存在临界静电不稳定性,表现为强正电势(Eij = +6.25 kJ/mol)和异常pKa值(−68.92 kT/e),提示电荷过饱和状态。PROSS与保守性分析进一步验证了R76Q作为进化兼容突变的最优性,可实现表面电荷的优化再分布。通过融合机器学习辅助预测和战略性二硫键工程,该框架实现了酶分子骨架的整体稳定性强化,其提升幅度超越了既往研究。
最终获得的首选突变体R76Q/T217C-G242C的性能验证了该策略的成功。该突变体表现出卓越的热稳定性,在55℃处理5分钟后仍保持71%残余活性。如表1所示,该性能优于已报道的ZEN水解酶:ZHD101和RmZHD在55℃下活性迅速丧失,CbZHD在45℃即失活,而本研究突变体保持强健功能。近期Xing等(2024)通过工程改造获得pH 4.2条件下比活力提升的M2突变体,但其热稳定性(50℃处理10分钟后残余活性40%)仍逊于本研究突变体在更严苛热胁迫下的表现。这使R76Q/T217C-G242C成为独具优势的强健生物催化剂,能够在工业加工相关的更宽温度和pH范围内保持活性。
本研究的另一显著成果是同步提升了酸稳定性——这一特性虽对发酵食品应用至关重要,却常被忽视。T217C-G242C二硫键的引入发挥了关键作用,使pH 5.0条件下的残余活性从I160Y的3%激增至100%。这证明战略性结构强化可同时赋予热稳定性和耐酸性。机制解析表明,这种双重稳定效应源于多重作用的协同:R76Q突变优化了全局静电环境,而T217C-G242C二硫键通过共价连接刚性化结构,限制构象柔性,从而同时抑制热解折叠和酸诱导变性——这与二硫键降低溶剂可及性、稳定催化位点的经典机制相符。
需要指出的是,从高性能实验室生物催化剂到获得批准的食品加工助剂,仍需开展进一步验证。解析复杂食品基质中降解产物的完整毒理学特征将是下一步研究的重点。尽管如此,本研究建立的多维优化框架为食品级酶的理性设计提供了通用而强大的技术蓝图。未来工作将聚焦于针对特定复杂食品体系定制此类工程化生物催化剂,并拓展其降解多种共存真菌毒素的能力。通过持续跨学科协作,这些进展有望为全球食品供应链安全保障提供可持续的酶学解决方案。
总结
综上所述,本研究通过创新性的混合计算设计策略,成功突破了玉米赤霉烯酮内酯水解酶在食品应用中关键的稳定性瓶颈。工程化改造的R76Q/T217C-G242C突变体在工业相关条件下展现出优异性能:40℃下半衰期延长121.8分钟,55℃处理5分钟后残余活性提升10倍,pH 5.0条件下耐酸性增强超过25倍。尤为关键的是,该突变体在霉变白酒酒糟这一复杂真实食品基质中,对天然存在的ZEN降解率达95.13%,凸显其实际应用潜力。本研究建立了一个稳健且可扩展的多维工程框架,协同整合了基于能量、静电作用、进化信息和结构强化的分析方法。该范式为稳健生物催化剂的理性设计提供了通用技术蓝图。尽管仍需在不同食品体系中进一步验证,但本研究不仅提供了高性能酶学解决方案,更构建了战略性工程框架,为食品工业实现更高效、可持续的真菌毒素防控开辟了新路径。






 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
