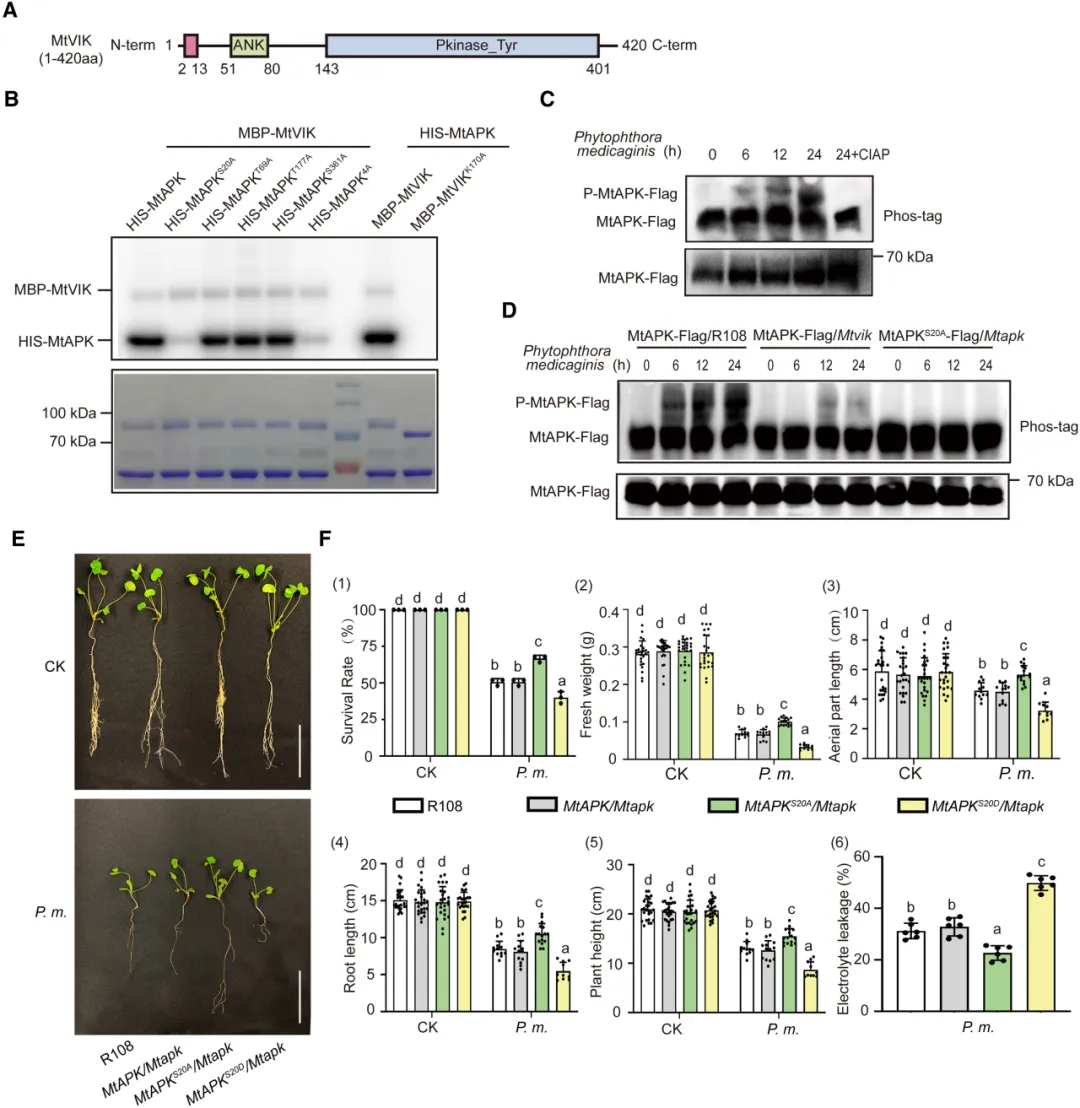
VIK是一种具有激酶活性的MAPKKK,因此MtVIK可能通过磷酸化下游目标蛋白,传递并放大刺激特异性信号到细胞中发挥磷酸化作用。MtVIK蛋白包含一个串联的Kyrin结构域和一个Pkinase Tyr结构域(图5A)。用丙氨酸(Ala)取代MtAPK的苏氨酸(Thr)和丝氨酸(Ser)残基产生磷酸化缺陷突变体。当Ser 20突变为Ala时,MtVIK对MtAPK的磷酸化作用失活。用Ala取代MtVIK的赖氨酸(Lys)残基,产生激酶死亡突变体(MBP-MtVIKK170A)。以未突变形式MBP-MtVIK作为对照进行体外磷酸化试验,结果表明,当MtVIK失去激酶活性时不能磷酸化MtAPK(图5B)。
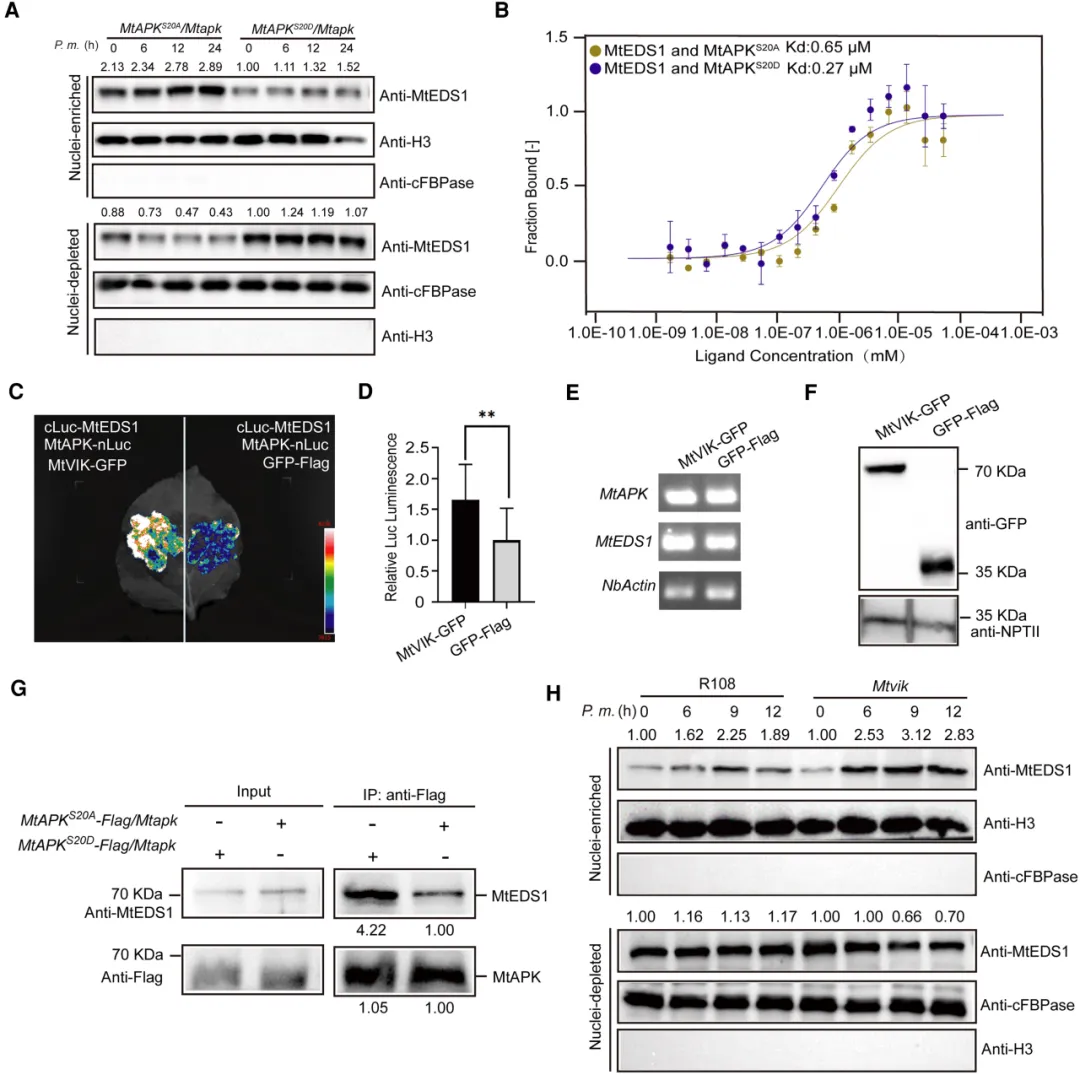
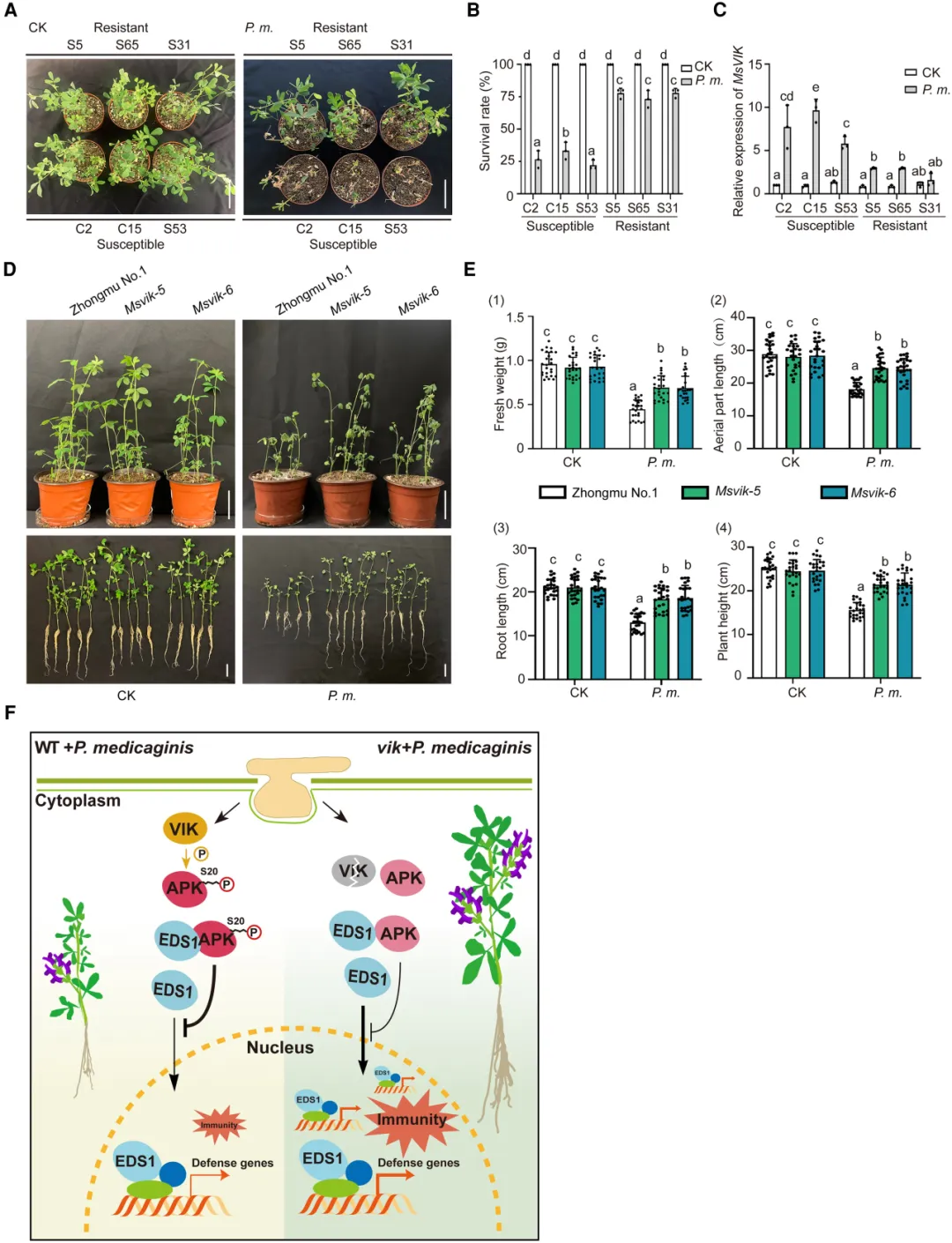
在Mtapk突变体中构建MtAPK自身启动子驱动的APK-FLAG回补植株(ProMtAPK:APK-FLAG/Mtapk),并检测其接种苜蓿疫霉菌前后MtAPK的磷酸化状态。Phos-Tag分析表明,MtAPK磷酸化水平的增加与接种时间的延长有关,用CIAP(小牛肠道碱性磷酸酶)处理后,磷酸化条带消失(图5C)。分别以野生型R108和Mtvik突变体为遗传背景获得表达MtAPK-Flag的转基因毛状根,与R108相比,Mtvik突变株的MtAPK磷酸化水平显著降低,与病原菌诱导MtVIK磷酸化MtAPK一致。以Mtapk突变体为遗传背景构建MtAPKS20A-Flag转基因毛状根(MtAPKS20A/Mtapk,将MtAPK第20位丝氨酸突变为丙氨酸),并观察到接种苜蓿疫霉菌后MtAPK磷酸化水平显著降低(图5D)。这些结果表明,MtVIK对MtAPKS20的磷酸化在抗苜蓿疫霉菌中起重要作用。此外,MtAPKS20A/Mtapk植株在接种苜蓿疫霉菌后各项生理指标(包括存活率、鲜重、地上部分长度、根长、株高和离子渗透率)显示,与野生型R108和MtAPKS20D/Mtapk(以Mtapk突变体为遗传背景构建MtAPKS20D-Flag转基因毛状根,将MtAPK第20位丝氨酸突变为天冬氨酸)相比,其抗性显著增强(图5E-F),说明MtAPKS20磷酸化有利于苜蓿疫霉菌感染。这些结果表明,苜蓿疫霉菌通过感染触发VIK磷酸化APK的Ser20来降低植物免疫。




 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
