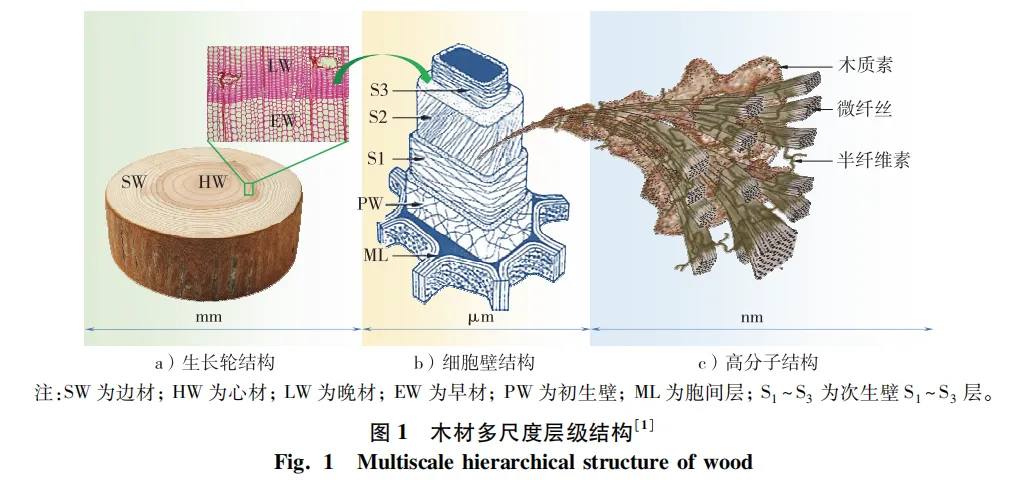
树木经亿万年自然选择,在常温常压下主要将碳、氢、氧化合物组装成复杂而巧妙的木质部层级结构,并实现了关键的结构功能,同时表现出独特的高强度和高韧性。在木材多尺度结构(图1)中,胞间层界面连接相邻2个细胞结构,微纤丝与木质素间通过半纤维素界面连接,微纤丝内部纤维素分子间通过分子间氢键界面连接。这些连接界面的共同特点是体积占比小,且强度和韧性比微纤丝、细胞壁或木材整体小1~3个数量级,因此,将这些连接界面称为木材弱界面。
“弱”表示界面自身力学性能与其连接的木材结构相比强度较小,但这并不表示弱界面在木材结构中没有积极作用。
相反,胞间层、半纤维素等连接弱界面具有适宜的界面结合强度,能够促使木材细胞壁、微纤丝等结构发生剪切滑移,增加木材塑性变形,导致裂纹尖端应力钝化和裂纹扩展偏转,提高了木材韧性,木材依此来应对外界较大的载荷刺激并确保其自身的结构安全。
此外,热处理、化学改性等改性技术可显著改善木材尺寸稳定性、生物耐久性,但改性材的韧性往往会有所降低,其主要原因之一是热处理和化学改性导致的弱界面特性发生了变化。
因此,研究木材分级结构中弱界面的物理化学结构及力学特性,揭示热处理和化学改性对多尺度弱界面的影响规律,并阐明弱界面对木材变形和韧性的影响机制,对木材及其改性材的物理力学性质研究具有重要意义。
1.1 木材细胞间的弱界面
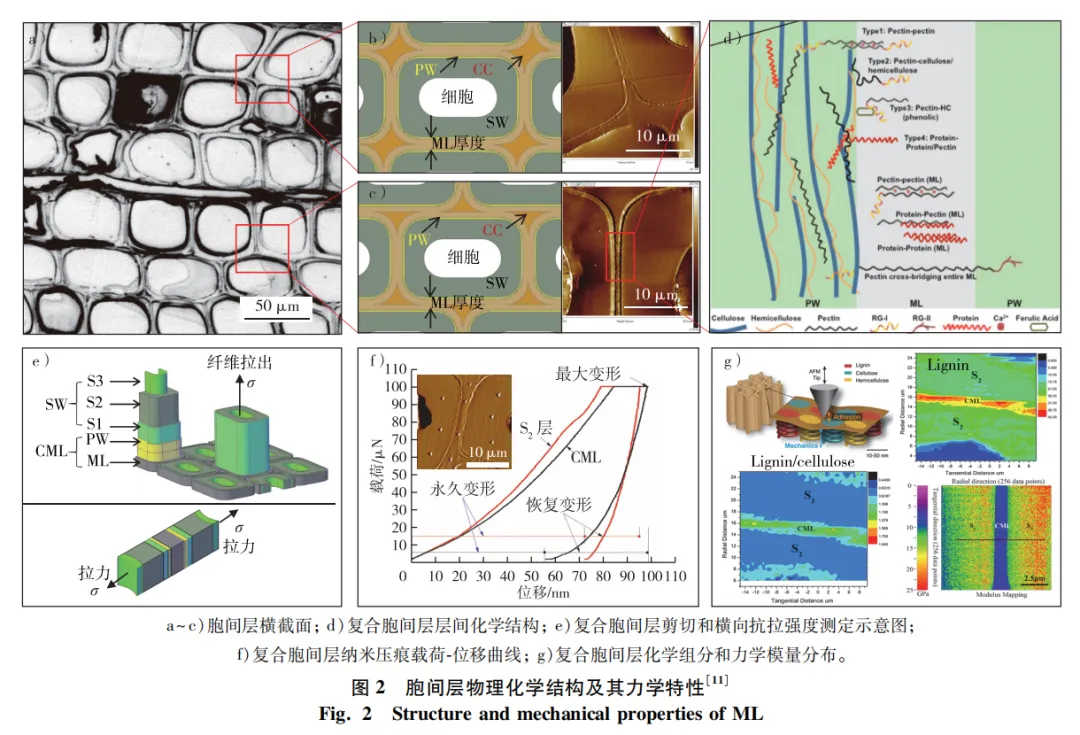
胞间层(ML)作为相邻细胞间的连接弱界面,对维持木材结构完整性至关重要,且有助于转移或分配施加到木材上的内外部载荷,确保木质部自身安全。胞间层弱界面的形状、尺寸和材料特性对研究木材结构及力学性质具有重要意义。胞间层具有网状几何结构,由连接相邻的2个细胞层状平面和2个以上细胞形成的细胞角隅(CC)组成,其几何形状可通过层状平面厚度和角隅形状来表征(图2a~c)。由于胞间层厚度较小,且很难将其与相邻的初生壁(PW)分离,因此通常使用复合胞间层(CML)几何参数来表示胞间层大小。如云杉管胞复合胞间层厚度为200~400 nm,而角隅部位的厚度高达1 200 nm,形状有内凹的三边形或四边形。胞间层是细胞质分裂过程中产生的细胞板,分裂初期细胞板是由果胶构成的多孔性网状结构(图2d),分裂后期在Ca2+离子作用下果胶的去酯化反应导致细胞板多孔性网状结构逐渐被填充和硬化,最后形成光滑的胞间层。成熟胞间层含有大量木质素和果胶(图2g),木质素质量分数可高达50%,远高于相邻次生壁中的木质素质量分数(约高20%)。
与次生壁相比,胞间层在强度和韧性上是一种相对较弱的材料,其抗拉强度为1~10 MPa,断裂韧性为0.1~0.3 kJ/m2,其强度和韧性都比正常木材低1~2个量级。目前,胞间层的力学性质主要借助原子力显微镜、纳米压痕仪等原位微区力学性能表征技术进行研究。例如,胞间层的压痕模量约为细胞壁S2层的一半,但其纵向压缩变形以及变形恢复能力均大于次生壁S2层(图2f)。虽然胞间层弱界面的物理、化学及力学性质方面研究取得了大量结果,但与次生壁研究相比依然存在不足。未来在胞间层弱界面结构解译方面,应从发育角度揭示胞间层弱界面的化学构成、组分含量和生物合成分子途径; 在胞间层弱界面力学性质研究方面,将胞间层从相邻细胞壁分离出来,直接测试其强度、应力-应变等力学性质,并构建胞间层弱界面微区结构与物理力学性能间的构效关系。
1.2 木材大分子间的弱界面
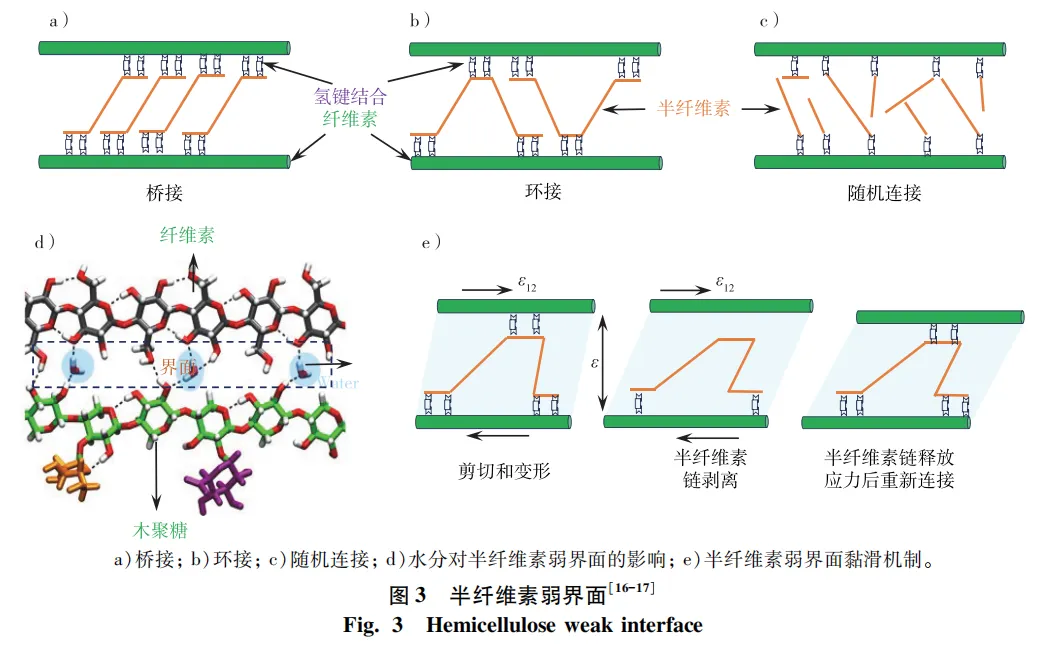
木材主要大分子间的交联结构一直是木材领域的研究热点,现已基本明确纤维素和木质素不直接相连,而是通过半纤维素连接。半纤维素与纤维素微纤丝表面可能的界面连接包括桥接(bridge)、环接(loop)和随机连接(random)这3种形式(图3a~c),且界面内可能的连接键包括分子间氢键、偶极-偶极相互作用等。此外,半纤维素与木质素主要通过静电吸附等物理作用以及共价键形式连接,其中木聚糖与低聚合度的木质素紧密相连,而葡甘露聚糖与高缩合度的木质素和纤维素紧密相连。但半纤维素与微纤丝表面、木质素结构的界面连接形式以及界面结合强度依然存在诸多未知及不确定性,有待进一步研究。
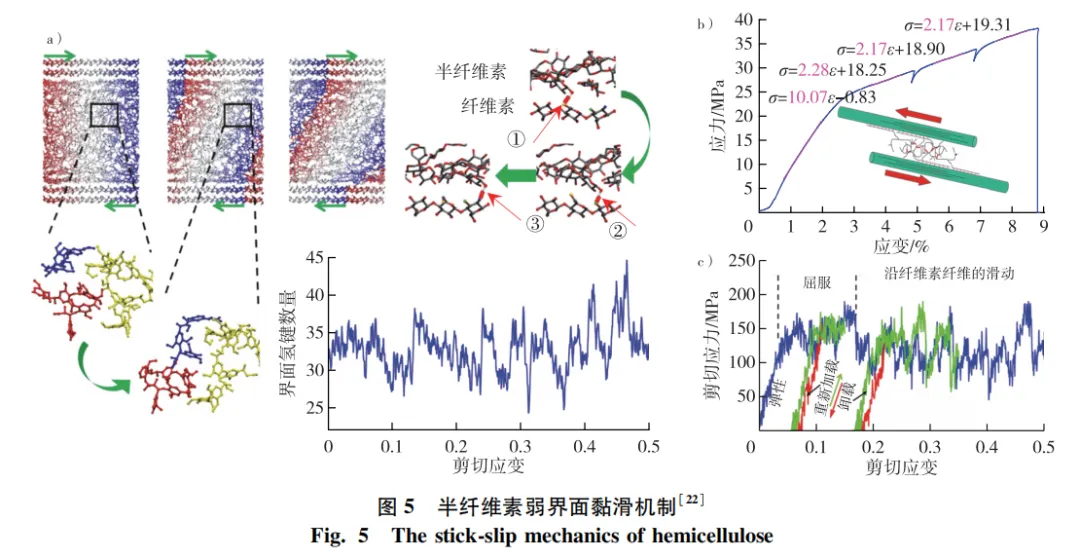
半纤维素作为纤维素微纤丝与木质素间的连接界面,其强度低,弹性模量约为20 MPa,较纤维素模量低3个数量级,但半纤维素增加了细胞壁的塑性变形。随着光谱设备与力学装置联用等表征技术的发展,大分子间的弱界面桥接作用和力学响应已成为木材构效关系基础研究领域的热点之一。研究结果表明:一方面,半纤维素弱界面促使纤维素微纤丝间产生剪切滑移,提高木材细胞壁的塑性变形能力和韧性; 另一方面,为木材提供一种“黏滑”机制,使得木材在应力去除后依然可以恢复到原来的强度(图3e)。
木材中的水分主要包括自由水和结合水,其中结合水主要存在于木材细胞壁内,决定木材物理力学性能。进入木材细胞壁的水分子优先与半纤维素结合,主要聚集在半纤维素弱界面区(图3d),只有小部分与微纤丝表面和木质素结合。水在半纤维素-纤维素界面上起着关键作用,水分进入细胞壁导致大分子间弱界面的界面连接形式发生变化,甚至解耦,界面结合强度发生变化,水分在木材大分子间起到一种“润滑剂”的作用,增加了细胞壁塑性变形,提高了细胞壁内的能量损失。动态红外研究结果表明:纤维素微纤丝表面与半纤维素连接的氢键不直接参与承载,但对湿木材的微纤丝间剪切滑移“黏滑”机制具有重要意义。此外,脱水引起大分子明显收缩后,半纤维素与纤维素更加紧密和不可逆地联系在一起,这些不可逆的纳米结构变化为解释和改善木基材料的性能提供了基础。因此,充分了解木材与水分的相互作用,解析水分进出细胞壁时大分子间弱界面的连接形式及界面结合强度变化,将对在分子尺度上阐明细胞壁脱水对木材力学性能的影响机制研究具有重要意义。
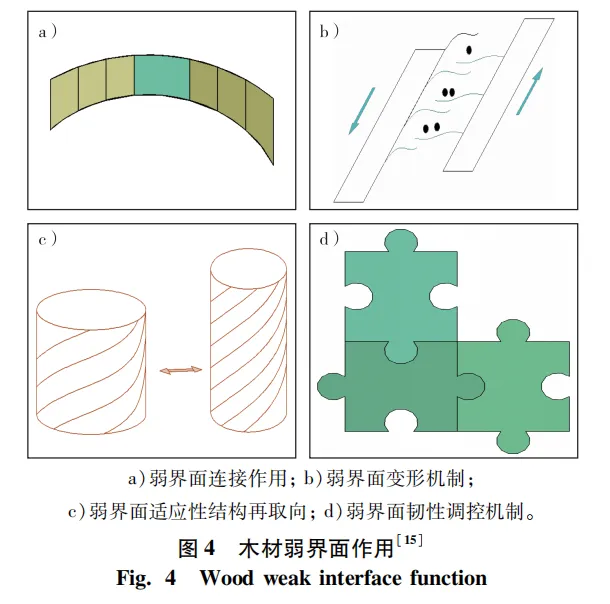
首先,弱界面在木材结构中起连接作用(图4),胞间层连接相邻2个细胞,半纤维素连接微纤丝与木质素,分子间氢键连接相邻纤维素大分子链。其次,弱界面促使木材细胞和微纤丝发生剪切滑移,使木材发生较大塑性变形。再者,弱界面允许细胞和微纤丝发生适应性结构再取向行为。适应性结构再取向是木材应对外部载荷变化时,细胞或微纤丝剪切滑移导致其取向发生变化,促使木材沿载荷方向力学性能发生强化。最后,弱界面有助于提高木材韧性,主要原因是弱界面引发的塑性变形使木材内部的裂纹尖端应力钝化和裂纹扩展发生偏转。
2.1 木材多尺度弱界面对木材变形的影响
光谱-力学联用、同步辐射光谱、分子动力学模拟等方法研究表明,顺纹拉伸时纤维素作为木材主要承载物质,而半纤维素弱界面及木质素基体主要起传递应力载荷的作用,且微纤丝间的剪切滑移受到半纤维素弱界面剪切阻力影响。Keckes等提出细胞壁“黏滑”剪切机制,即当剪切应力超过一定水平时,微纤丝发生剪切滑移,类似于金属材料的位错滑移,从而在较大的滑动距离内提供内聚行为。当应力释放时,微纤丝又被锁定在变形位置,产生不可恢复的塑性变形,但木材不会累积损伤或失去刚度。“黏滑”剪切机制可能是由半纤维素弱界面引起的(图5),半纤维素与微纤丝间的横向结合需要部分半纤维素形成氢键并与微纤丝在一定距离上对齐,从而在界面上形成不连续的半纤维素桥。当界面处的剪应力超过临界剪应力时,半纤维素弱界面可能会从其中一侧微纤丝上脱离,释放了一些桥接强度,并允许微纤丝间发生剪切变形。
2.2 木材多尺度弱界面对木材适应性结构取向的影响
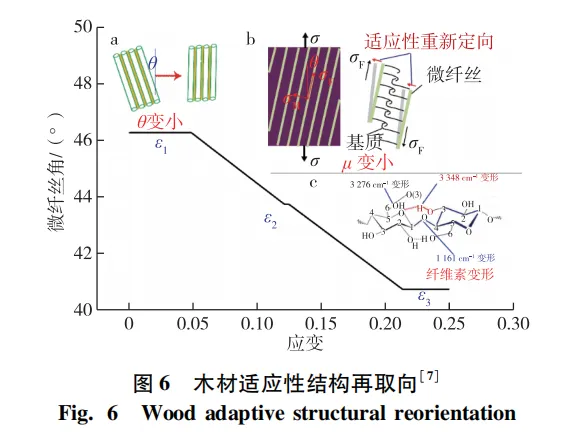
木材受到外载荷时,弱界面促使其连接结构发生剪切滑移,导致木材细胞和纤维素微纤丝取向发生变化,木材的这种响应机制称为适应性结构再取向。适应性结构再取向是木材、骨骼和角质层等生物材料所特有的,是生物材料应对外部载荷一种被动的应变响应机制,为其提供了一种特殊的变形机制。木材适应性结构再取向主要包括胞间层弱界面促使的木材细胞适应性结构再取向(图6a)和半纤维素弱界面促使的微纤丝适应性结构再取向(图6b)。木材细胞和微纤丝再取向过程中细胞之间和微纤丝之间都会发生剪切滑移,并且剪切滑移变形大于细胞壁和微纤丝自身的伸长变形,从而为木材提供了较大塑性变形能力,也为木材提供了一种增韧机制,以提高木材的断裂韧性和能量耗散能力。
木材适应性结构再取向主要受弱界面性质、微纤丝角、含水率等因素影响。胞间层的塑性变形决定了木材细胞之间的剪切滑移变形大小,从而成为影响细胞适应性结构再取向的关键因素。半纤维素弱界面的桥接作用及力学性质是影响微纤丝适应性结构取向的关键因素。再者,木材初始微纤丝角主要决定了微纤丝的受力方向及大小,影响微纤丝的剪切滑移,成为影响微纤丝适应性结构再取向的另一个关键因素。最后,水分易导致弱界面结构和厚度发生变化,水分作用下胞间层弱界面的塑性变形增加,导致细胞和微纤丝间的剪切滑移变形增加,从而影响木材适应性结构再取向,但以上因素各自多大程度上影响木材结构适应性再取向目前还不清楚。此外,建立细胞和微纤丝的取向变化与应变的量化关系,将有助于揭示弱界面对木材适应性结构再取向的影响机制。
2.3 弱界面对木材韧性和强度的影响
韧性表示木材对裂纹或裂纹状缺陷扩展的抵抗力,裂纹会引起木材局部应力集中,导致裂纹尖端周围材料失效,且裂纹不可避免地会进行扩展,最终导致材料失效。木材韧性主要通过断裂韧性、冲击强度和塑性变形吸收能来表示,韧性越好,木材发生脆性断裂的可能性越低。木材属于各向异性材料,裂纹垂直木材纹理方向扩展的韧性相当高,约为104 J/m2。这与工程铝合金或中碳钢韧性相当,但裂纹平行木材纹理扩展的韧性较低,比垂直纹理方向扩展的木材韧性低2个数量级。
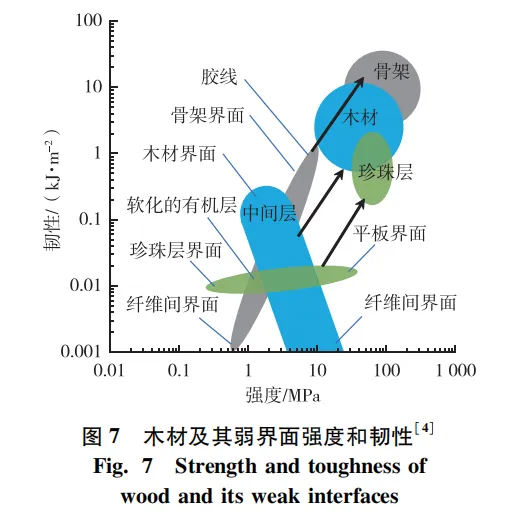
强度和韧性往往是一对矛盾力学关系,强度提高往往会降低其韧性,导致材料脆性增加,但木材很好地兼容了强度和韧性,属于高强韧化材料。微纤丝剪切滑移导致的细胞壁塑性变形、纤维拨出以及微纤丝和基体自身的断裂破坏都会影响木材韧性,但微纤丝和基体自身破坏对木材整体韧性的贡献很小(10~100 J/m2)。这是因为与大多数人造复合材料不同,木材结构中包含了不同尺度的弱界面(图7),能够促使微纤丝发生较大的剪切滑移,甚至纤维拨出,从而激发木材能量吸收机制; 此外,弱界面导致的塑性变形能够降低裂纹尖端的应力集中,使得裂纹扩展受到阻碍,增加木材韧性。在木材结构中,裂纹在弱界面处被钝化或偏离,裂纹尖端半径的增大导致应力集中系数降低,从而阻止裂纹的扩展。只有界面强度与木材性能相适应时,这种增韧机制才会起作用,既不能太弱,也不能太强。Cook等首次提出了弱界面的裂纹钝化和偏转机制,当弱界面强度约为周围材料的1/5时,弱界面就会促使裂纹被钝化或偏转,从而刺激能量吸收。
3.1 热处理改性对木材弱界面及韧性的影响
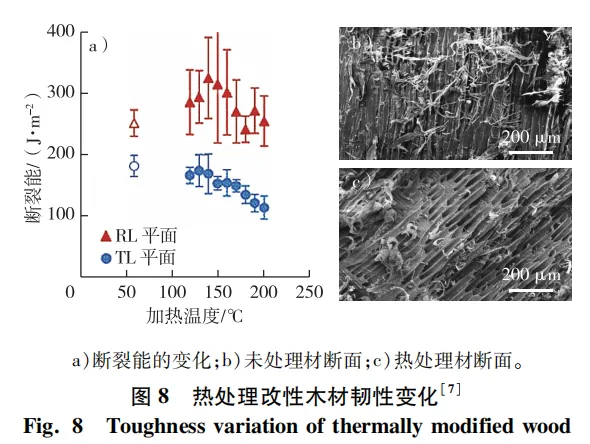
以饱和水蒸气、氮气、热油及真空作为加热介质的木材热处理技术显著提升了木材尺寸稳定性、耐腐性以及耐候性,但热处理木材化学结构改变和质量损失导致其部分力学性能下降,其中韧性损失尤为明显,限制了其作为结构材的使用范围(图8)。热处理介质、温度以及时间是影响热处理材韧性大小的重要因素。例如:160~170 ℃处理24 h,木材断裂韧性降低50%~80%; 180 ℃处理6 h,欧洲赤松和辐射松的冲击韧性分别下降了56%和80%; 200 ℃氮气介质加热处理12 h,木材脆性比未处理材大4倍; 170~230 ℃的高温蒸汽热处理后,马尾松的脆性增加 33.64%~89.01%。
热处理过程中半纤维素弱界面最不稳定,半纤维素弱界面和纤维素间通过大量氢键连接,氢键在剪切应力作用下的断裂和在新位置上的重新结合赋予了木材一定的分子流动性和塑性,且木材90%的断裂能量被塑性变形吸收。试验表明,热处理后韧性的降低主要源于塑性延展性的降低,热处理后裂纹更易发生,裂纹传播阻力降低。此外,热处理也会导致大分子之间发生交联反应,如热处理过程中细胞壁内成分发生酯化反应,酯基与木质素相连。Salmén等利用动态红外光谱研究也发现,热处理后半纤维素和木质素的力学贡献增加,表明热处理后木材细胞壁大分子间的交联明显增强。热处理木材细胞壁的流动性可以通过细胞壁黏弹性来衡量。动态纳米压痕研究结果表明,热处理后木材细胞壁和胞间层的损耗模量和损耗因子降低,细胞壁黏性降低,延伸性下降,导致热处理材的韧性降低。目前大部分学者认为,半纤维素的降解是热处理木材韧性降低的主要原因,但该观点与多种试验证据也存在不相符的情况。如化学法脱除半纤维素后,木材细胞壁的黏性和塑性反而提高。与湿热处理相比,干燥条件下热处理后木材的半纤维素降解更少,但脆性更大。因此,在分子水平上,热处理木材韧性的变化存在半纤维素降解和大分子交联增加这两种可能的机制,都可能使得热处理材细胞壁塑性减小,裂纹易产生和扩展,从而导致热处理材韧性降低、脆性增加。
3.2 化学改性对木材弱界面及韧性的影响
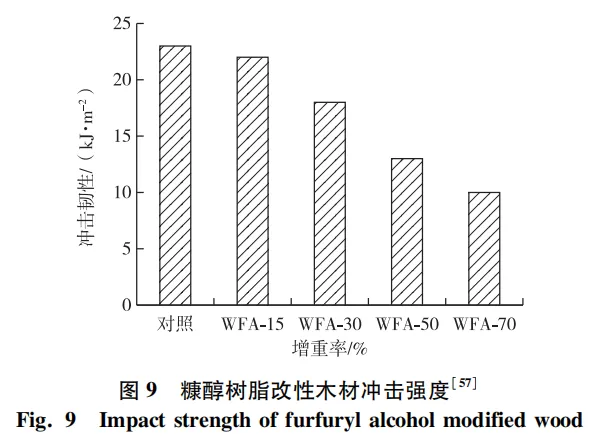
化学改性主要是通过化学方法来改变细胞壁特性或填充细胞腔,旨在改善木材尺寸稳定性、机械性能和抗生物降解性。细胞壁改性是采用乙烯等活性低分子量单体,或糠醛等低分子量树脂与细胞壁组分发生反应,从而改变细胞壁成分。细胞腔填充则是通过化学物质沉积在细胞腔中,并阻塞细胞物理通道。但与热处理改性类似,化学改性也会导致木材的韧性降低,且韧性降低程度与改性剂类型、增重百分比、改性剂分子结构、处理温度和时间等都有关系。例如:乙酰化改性苏格兰松木材的断裂韧性降低36%~50%; 三聚氰胺甲醛树脂改性木材的冲击韧性降低20%~90%; 糠醇改性木材的冲击强度降低4%~57%(图9),且随着糠醇浓度增加,改性材的冲击强度降低程度增加。
化学改性木材韧性降低、脆性增加的可能原因有:①改性树脂脆性较大导致的。如糠醇改性材韧性的降低被认为是由脆性糠醇树脂导致的,可通过环氧植物油降低糠醇脆性,从而减小糠醇改性材的韧性损失。②改性树脂酸碱特性,甚至固化温度都会导致半纤维素弱界面降解,破坏细胞壁的完整性,影响裂纹扩展,降低木材韧性。③树脂可能与木材细胞壁主成分表面发生化学交联反应,从而改变大分子间的弱界面强度,生成的高刚性热固性聚合物限制细胞壁分子链的流动性和细胞壁剪切滑移,导致木材韧性降低。但以上因素究竟各自如何和多大程度上导致改性材韧性降低目前鲜见研究。
与细胞壁、微纤丝等起强度作用的木材结构模块相比,木材弱界面研究并不充分,其主要原因是弱界面尺度小,精细化的结构与性能表征难以实现。此外,现有热处理、细胞壁化学改性等绝大多数木材改性方法都会引起木材弱界面特性结构变化,导致改性材脆性增加,限制改性材的使用范围。因此,围绕木材弱相结构未来可重点开展以下研究:
1)在木材多尺度弱界面研究方面。在细胞壁尺度,开展胞间层弱界面微区结构与物理力学性能间构效关系的研究,解译胞间层弱界面对细胞壁多层结构的应力分配特性。在分子尺度,开展分子间弱界面连接形式及界面结合强度表征研究,揭示弱界面连接形式、水分等对弱界面结合强度的影响机制。
2)在木材变形与韧性机制研究方面。重点开展细胞壁和分子尺度上的木材多尺度结构变形表征研究,揭示微纤丝角及大分子间的弱界面对细胞壁塑性变形以及微纤丝再取向的影响机制。研究弱界面激发的细胞壁塑性变形以及“黏滑”机制,阐明弱界面对木材韧性的影响。
3)在改性木材韧性研究方面。开展热处理、细胞壁化学改性等改性技术对胞间层弱界面构效关系的影响研究,分析改性处理对大分子间的弱界面结合强度以及大分子间剪切滑移变化的影响。开展改性材多尺度弱界面变化对细胞壁塑性变形以及裂纹偏转的影响研究,阐明热处理和化学改性对韧性降低的影响机制。





 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
