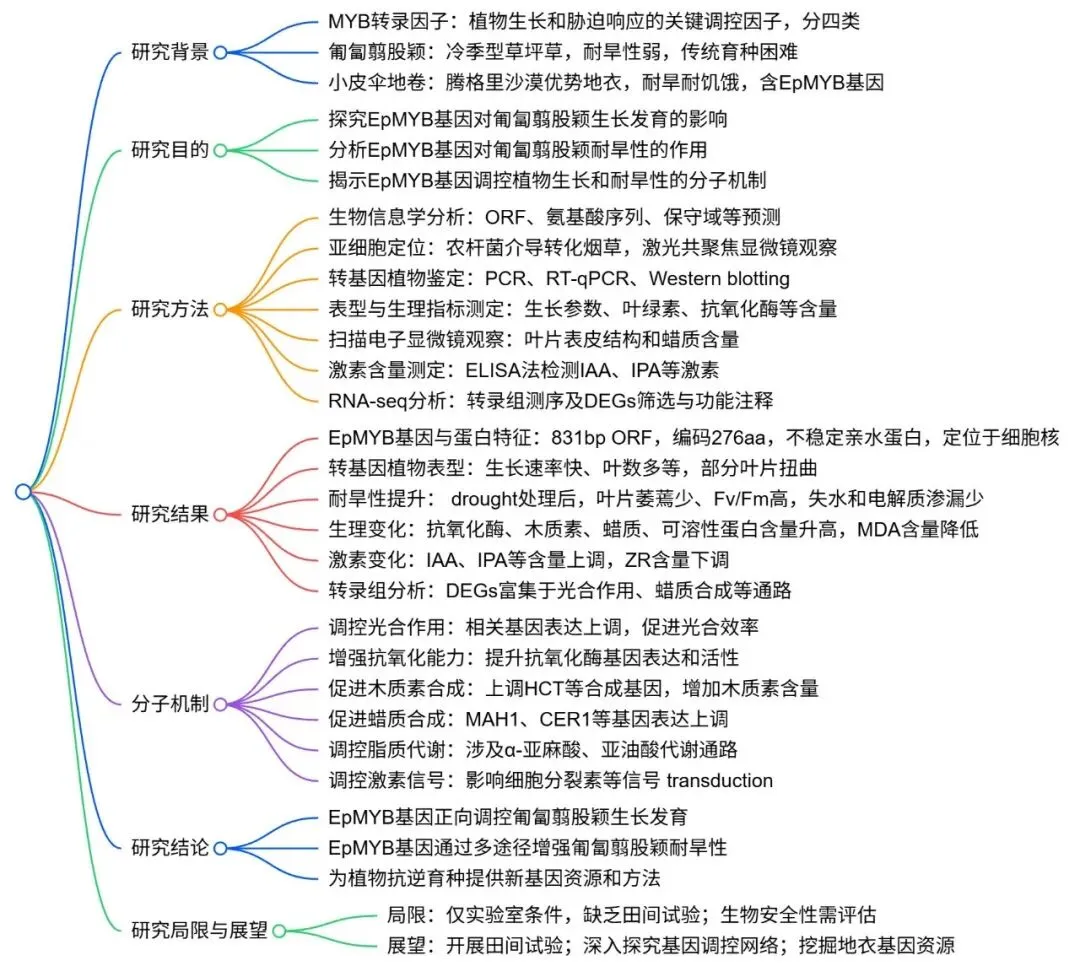
3.1EpMYB的生物信息学分析
基因结构:EpMYB基因开放阅读框为831 bp,编码276个氨基酸。
蛋白结构域:在92至134位氨基酸之间存在一个MYB-DNA结合域。
理化性质:EpMYB蛋白分子量为30.92 kDa,理论等电点为9.65,属不稳定亲水蛋白。
二级结构:蛋白主要由α-螺旋(27.17%)和无规卷曲(57.97%)构成。
信号肽预测:EpMYB蛋白不含信号肽。
EpMYB(XM_007804716.1)基因含有一个831 bp的开放阅读框,编码276个氨基酸。通过NCBI网站进行的保守结构和性质预测分析发现,该蛋白在第92至134位氨基酸之间存在一个MYB-DNA结合域,对应于MYB结合位点(Fig. 1 A)。EpMYB的分子式为C₁₃₄₇H₂₁₁₈N₄₁₀O₄₁₁S₉,总分子量为30.92 kDa,理论等电点为9.65。其正、负电荷残基数分别为39和29。不稳定性指数(II)为79.98,亲水性总平均值(GRAVY)为-0.924,表明EpMYB是一种不稳定的亲水蛋白。二级和三级结构预测显示,EpMYB蛋白主要由α-螺旋(27.17%)和无规卷曲(57.97%)组成,同时含有少量延伸链(10.51%)和β-转角(4.35%)(Fig. 1 B, C)。此外,SignalP 6.0预测表明EpMYB蛋白不含信号肽(Fig. 1 D)。
图1.EpMYB的生物信息学分析。EpMYB蛋白的预测结构域(A),EpMYB蛋白的二级结构预测(B),EpMYB蛋白的三级结构预测(C),以及EpMYB蛋白的预测信号肽(D)。
3.2 亚细胞定位
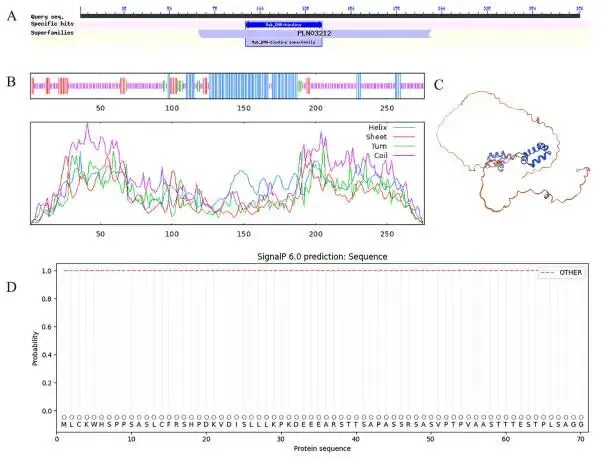
蛋白定位:EpMYB蛋白定位于细胞核中。
定位验证:DAPI染色进一步证实了其细胞核定位。
将3302Y空载体和3302Y-EpMYB重组载体分别通过农杆菌介导法转入烟草叶片。经过两天的暗培养后,通过共聚焦荧光显微镜观察发现,细胞核中有强烈的荧光信号。此外,利用4’, 6-二脒基-2-苯基吲哚(DAPI)对细胞核进行染色,结果进一步证实EpMYB蛋白定位于细胞核中(Fig. 2)。
图2.EpMYB蛋白的亚细胞定位。Free YFP,空载体对照。EpMYB-YFP,EpMYB-YFP融合蛋白。细胞核经4’, 6-二脒基-2-苯基吲哚(DAPI)染色,以确认EpMYB蛋白在细胞核中的位置。标尺 = 25 μm。
3.3 转EpMYB基因植物的获得
PCR结果证实EpMYB基因已整合到转基因匍匐翦股颖的基因组中(Fig. S1)。实时定量聚合酶链式反应(RT-qPCR)结果表明,EpMYB基因在转基因(TG)植株中正常表达(Fig. S2)。Western blotting检测证明,TG植株中成功产生了EpMYB蛋白(Fig. S3)。
3.4 野生型与转基因匍匐翦股颖的表型及内部结构特征
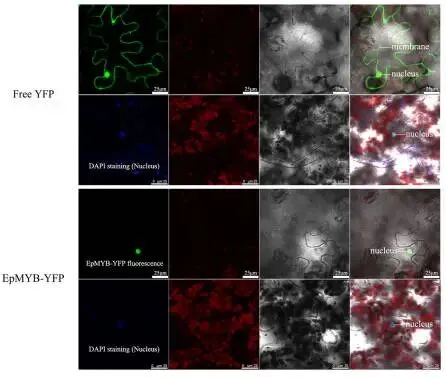
生长速率:TG植株生长速率更快,地上部和根系鲜重分别增加44.8%和41.6%。
农艺性状:TG植株最大叶长、单分蘖叶数和节间长度分别增加25.6%、3.0倍和35.1%。
叶片表型:部分叶片表现出严重扭曲。
生理指标:TG植株叶绿素含量显著提高9.4%。
表皮结构:TG植株叶片气孔周围蜡质含量增加且形态改变。
基因表达:蜡质合成基因MAH1在TG植株中表达量显著上调2.5倍。
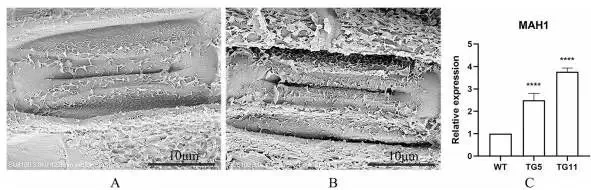
在相同条件下培养12周后,与野生型(WT)植株相比,TG植株表现出更快的生长速率(Fig. 3 A, B, C),其地上部鲜重和根系鲜重分别增加了44.8%和41.6%(Fig. 3 G, H)。同时,TG植株的最大叶长增加了25.6%(Fig. 3 D, I),单分蘖叶数显著增加了3.0倍(Fig. 3 F, J),节间也更长(增加了35.1%)(Fig. 3 K)。部分叶片出现严重扭曲现象(Fig. 3 E)。此外,TG植株的叶绿素含量比WT植株显著高9.4%(Fig. 3 L)。扫描电子显微镜观察发现,TG植株叶片气孔周围的蜡质含量增加,且形态发生改变(Fig. 4 A, B)。蜡质合成基因MAH1的表达水平在TG植株中比WT植株显著高2.5倍,表明EpMYB基因显著促进了TG植株的蜡质合成(Fig. 4 C)。
图3. 植株的表型特征和叶绿素含量。A. 分别在相同条件下于培养基中生长的野生型(WT)和转基因(TG)植株。B. 分别在土壤(营养土:蛭石:珍珠岩=1:1:1)中生长并在成熟后拍摄的WT和TG植株。C. 代表性的WT和TG匍匐翦股颖植株的根系。D, E. 代表性的WT和TG匍匐翦股颖植株的正常叶片(D)和扭曲叶片(E)。F. 来自WT和TG株系的代表性单分蘖表型。G-L. WT和TG代表性植株在地上部鲜重(G)、根系鲜重(H)、最大叶长(I)、单分蘖叶数(J)、节间长度(K)和叶绿素含量(L)方面差异的统计分析。数据为三次生物学重复的平均值±标准差(SD)。与WT相比,TG株系的显著性差异通过学生t检验用星号(ns, *, *, 或)表示,分别对应ns, P < 0.05, 0.01, 0.001或0.0001。
图4. 野生型(WT)对照和转EpMYB基因(TG)匍匐翦股颖株系的蜡质合成。WT(A)和TG(B)叶片气孔的扫描电子显微镜图像。C. WT和TG植株中蜡质合成基因MAH1的相对表达量。MAH1,中链烷烃羟化酶。数据为四次生物学重复的平均值±标准差(SD)。与WT相比,TG株系的显著性差异通过学生t检验用星号(ns, *, *, 或)表示,分别对应ns, P < 0.05, 0.01, 0.001或0.0001。
3.5 转基因植物通过提高抗氧化酶活性和渗透调节能力增强抗旱性
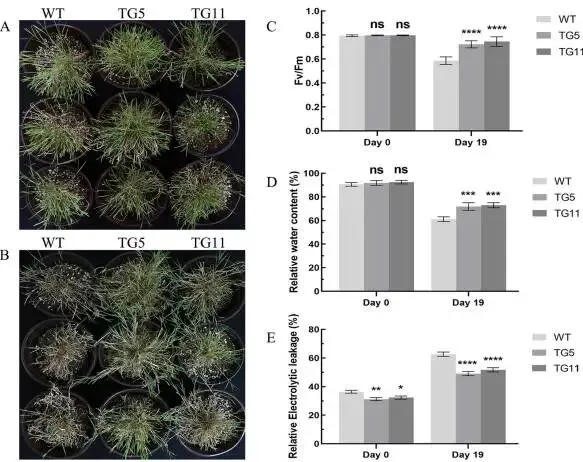
抗旱表型:干旱19天后,TG植株大部分保持绿色,而WT植株多数萎蔫变黄。
光合效率:TG植株的Fv/Fm值显著高于WT植株(高25.3%)。
水分状况:TG植株失水更少(少18.5%),电解质渗漏更低(低32.8%)。
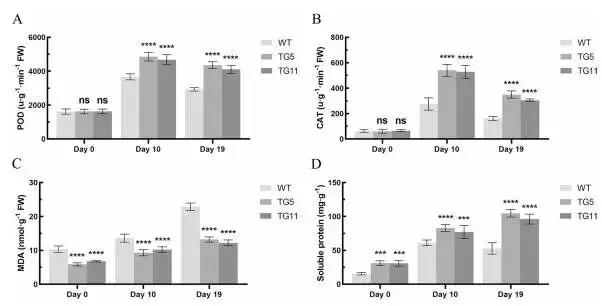
抗氧化酶:干旱处理后,TG植株的POD和CAT含量显著高于WT植株。
膜脂过氧化:TG植株的MDA含量显著低于WT植株。
渗透调节物:TG植株的可溶性蛋白含量在干旱胁迫下持续增加并显著高于WT植株。
对WT和TG植株进行自然干旱处理(Fig. 5 A)。在干旱处理第19天,大部分WT植株叶片萎蔫变黄,而TG植株仅有少数叶片变黄,大部分保持绿色(Fig. 5 B)。TG植株的Fv/Fm值比WT植株显著高25.3%(Fig. 5 C),表明EpMYB基因增强了TG植株的抗旱性。生理测量显示,经过19天的干旱处理,TG植株失水比WT植株少18.5%(Fig. 5 D),电解质渗漏低32.8%(Fig. 5 E),表明其因干旱胁迫造成的膜损伤较轻。在干旱处理10天和19天后,过氧化物酶(POD)和过氧化氢酶(CAT)的含量在WT和TG植株中均呈先增后降的趋势,但TG植株的POD(分别高30.1%和44.3%)和CAT(分别高94.1%和101.9%)水平显著高于WT植株(Fig. 6 A, B)。丙二醛(MDA)含量随时间在两类植株中均增加,但TG植株的MDA水平显著低于WT植株(分别低28.3%和44.5%)(Fig. 6 C),表明EpMYB通过增强抗氧化酶活性减轻了干旱诱导的损伤。WT植株的可溶性蛋白含量先增后降,而TG植株则持续增加,并显著高于WT植株(分别高30.6%和90.1%)(Fig. 6 D)。这些结果表明,EpMYB基因通过改善渗透调节能力来增强匍匐翦股颖的抗旱性。
图5. 野生型(WT)和转基因(TG)植株的干旱处理。WT和TG植株在干旱处理第0天(A)和第19天(B)的形态比较。在干旱处理0天和19天测量的Fv/Fm(C)、相对含水量(D)和相对电解质渗漏(E)。数据为三次生物学重复的平均值±标准差(SD)。与WT相比,TG株系的显著性差异通过学生t检验用星号(ns, *, *, 或)表示,分别对应ns, P < 0.05, 0.01, 0.001或0.0001。
图6. 野生型(WT)对照和转EpMYB基因(TG)匍匐翦股颖株系对干旱胁迫的生理响应和分子分析。在干旱处理第0、10和19天测量的WT和TG植株的生理指标,包括过氧化物酶(POD)(A)、过氧化氢酶(CAT)(B)、丙二醛(MDA)(C)、可溶性蛋白(D)。数据为三次生物学重复的平均值±标准差(SD)。与WT相比,TG株系的显著性差异通过学生t检验用星号(ns, *, *, 或)表示,分别对应ns, P < 0.05, 0.01, 0.001或0.0001。
3.6 转基因植物木质素含量增加
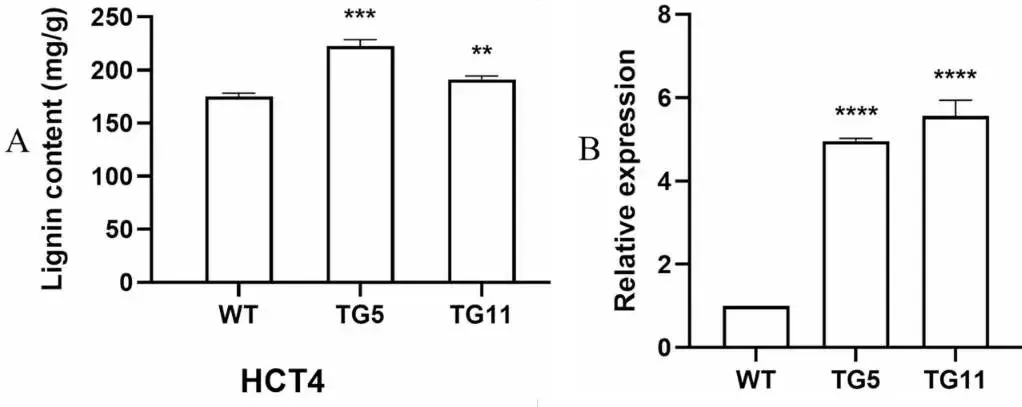
结果显示,与WT植株相比,两个转基因株系地上部的总木质素含量分别增加了25.2%和9.1%(Fig. 7 A)。此外,与WT植株相比,木质素合成基因HCT4在TG植株中的表达水平显著高4.2倍(Fig. 7 B),表明EpMYB基因促进了TG植株的木质素合成。
图7. 野生型(WT)对照和转EpMYB基因(TG)匍匐翦股颖株系的木质素。A. WT对照和TG植株的木质素含量。数据为三次生物学重复的平均值。B. WT和TG植株中木质素合成基因HCT4的相对表达量。HCT4,羟基肉桂酰转移酶4。数据为四次生物学重复的平均值±标准差(SD)。与WT相比,TG株系的显著性差异通过学生t检验用星号(ns, *, *, 或)表示,分别对应ns, P < 0.05, 0.01, 0.001或0.0001。
3.7 转基因匍匐翦股颖的激素变化
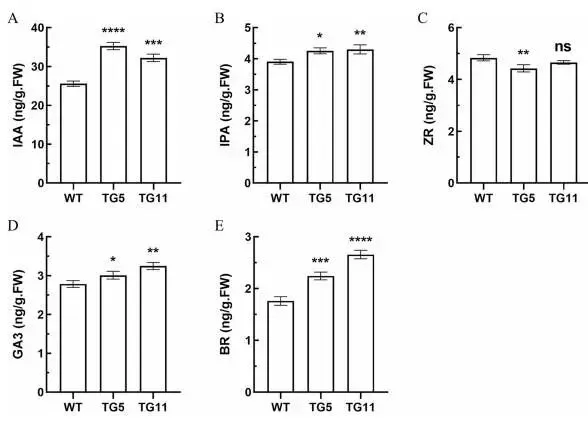
与WT植株相比,TG植株中的IAA(31.9%)(Fig. 8 A)、IPA(9.6%)(Fig. 8 B)、GA3(13.7%)(Fig. 8 D)和BR(40.1%)(Fig. 8 E)水平均上调,其中IAA和BR的增幅尤为显著。相反,ZR含量在TG植株中降低了6.1%(Fig. 8 C)。这些结果表明,EpMYB转录因子影响多种激素的协同调控,进而影响植物生长和对非生物胁迫的响应。
图8. 野生型(WT)和转基因(TG)植株地上部的激素含量。生长素(IAA)含量(A)。细胞分裂素异戊烯腺苷(IPA)含量(B)。细胞分裂素反式玉米素核苷(ZR)含量(C)。赤霉素3(GA3)含量(D)。油菜素内酯(BR)含量(E)。单位为ng/g·FW。数据为三次生物学重复的平均值±标准差(SD)。与WT相比,TG株系的显著性差异通过学生t检验用星号(ns, *, *, 或)表示,分别对应ns, P < 0.05, 0.01, 0.001或0.0001。
3.8 转录组测序结果
数据质量:12个样本共产生91.90 Gb的高质量数据,Q30介于94.19%-95.04%。
样本相关性:生物学重复样本间相关性高,表明实验可重复性好。
主成分分析:PCA分析显示不同处理组之间存在明显分离,组内重复性好。
基因注释:共获得54,408个unigene,其中36,741个得到注释。
差异基因统计:在TG-vs-WT、WT(干旱处理)-vs-WT和TG(干旱处理)-vs-TG比较组中分别鉴定出3,530、14,907和7,354个差异表达基因(DEGs)。
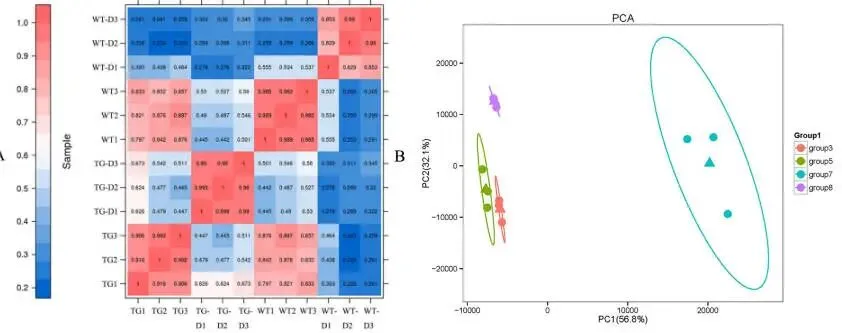
对12个样本进行转录组测序后,共产生91.90 Gb的清洁数据,每个样本的数据量为6.37 Gb。12个样本的Q30值范围在94.19%至95.04%之间,表明转录组数据的可靠性和准确性均较高(Table 1)。生物学重复间的相关性分析显示,重复样本间的相关性较高(Fig. 9 A),表明鉴定出的差异表达基因可靠性高。此外,主成分分析(PCA)结果揭示了清晰的聚类模式:重复样本紧密聚类,表现出高重复性,而不同组别之间则明显分离(Fig. 9 B)。组装后共获得54,408个unigene,其中23,252个长度超过1 kb。通过与多个数据库比对,共注释了36,741个unigene。在TG-vs-WT比较组中鉴定出3,530个DEGs(1,949个上调,1,581个下调);在WT(干旱处理)-vs-WT组中鉴定出14,907个DEGs(7,235个上调,7,672个下调);在TG(干旱处理)-vs-TG组中鉴定出7,354个DEGs(4,029个上调,3,325个下调)。
图9. 12个样本的相关性图和PCA散点图。A. 相关性图。B. PCA散点图。横纵坐标分别代表第一和第二主成分,括号中的数字表示各主成分的贡献度。三角形代表各组的平均坐标(中心点),椭圆框表示置信区间。不同分类标准用颜色区分。Group3,TG组。Group5,WT组。Group7,WT-D组。Group8,TG-D组。WT-D,干旱处理后的野生型对照;TG-D,干旱处理后的转基因植株。
3.9 TG-vs-WT组差异表达基因的GO和KEGG分析
上调GO富集:富集于光合作用、氧化应激反应、脂质代谢过程等。
下调GO富集:富集于mRNA转录、跨膜运输、生长素激活信号通路等。
KEGG通路富集:主要富集于光合作用、角质、栓质和蜡质生物合成、α-亚麻酸代谢等。
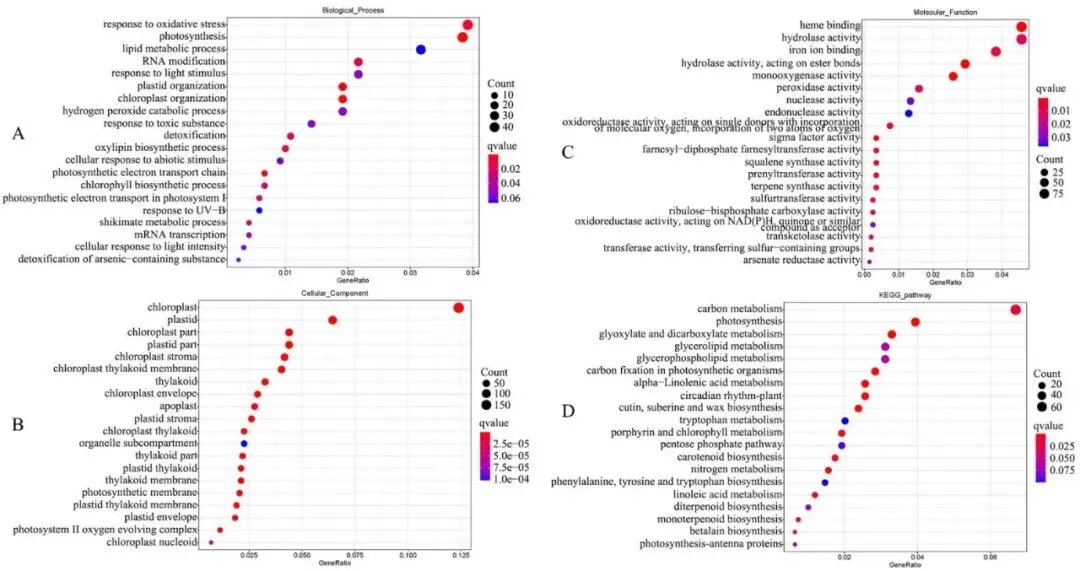
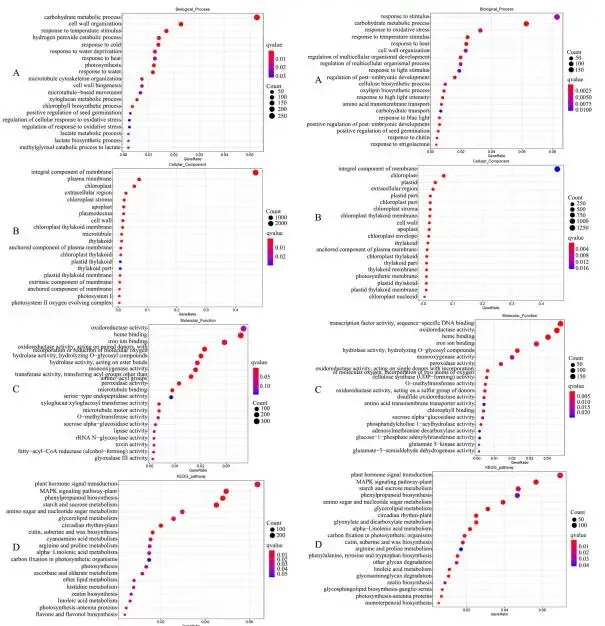
基于GO分析,上调的DEGs主要富集的生物学过程包括光合作用、光合电子传递链、氧化应激反应、脂质代谢过程等;下调的DEGs主要富集的生物学过程包括mRNA转录、莽草酸代谢过程、跨膜运输、生长素激活信号通路等(Fig. 10 A)。上调DEGs富集的细胞组分主要为叶绿体、类囊体膜等;下调DEGs富集的细胞组分主要为膜的组成部分、植物型液泡等(Fig. 10 B)。上调DEGs富集的分子功能包括水解酶活性、氧化还原酶活性、抗氧化活性等;下调DEGs富集的分子功能包括跨膜转运蛋白活性、铁离子结合等(Fig. 10 C)。KEGG分析显示,DEGs主要参与光合作用、角质、栓质和蜡质生物合成、α-亚麻酸代谢、亚油酸代谢、类胡萝卜素生物合成、甘油酯代谢、玉米素生物合成和类黄酮生物合成等通路(Fig. 10 D)。
图10. 野生型(WT)和转基因(TG)植株间基因表达的全局分析。A. TG-vs-WT组中所有注释的差异表达基因(DEGs)的基因本体论(GO)富集的生物学过程分析。B. TG-vs-WT组中所有注释DEGs的GO富集的细胞组分分析。C. TG-vs-WT组中所有注释DEGs的GO富集的分子功能分析。TG-vs-WT,转基因植株对野生型植株。根据p值选择了20个最显著的GO类别。D. TG-vs-WT组中注释DEGs的京都基因与基因组百科全书(KEGG)分析。TG-vs-WT,转基因植株对野生型植株。根据q值选择了20个最显著的KEGG通路。
3.10 WT(干旱处理)-vs-WT组差异表达基因的GO和KEGG分析
上调GO富集:富集于非生物刺激响应、缺水响应、L-脯氨酸生物合成过程等。
下调GO富集:富集于光合作用、细胞壁组织、碳水化合物代谢过程等。
KEGG通路富集:主要富集于角质、栓质和蜡质生物合成、苯丙烷类生物合成、淀粉和蔗糖代谢等。
基于GO分析,上调的DEGs主要富集的生物学过程包括对非生物刺激的响应、对缺水的响应、L-脯氨酸生物合成过程、自噬等;下调的DEGs主要富集的生物学过程包括光合作用、细胞壁组织、碳水化合物代谢过程、过氧化氢分解过程等(Fig. 11 A)。上调DEGs富集的细胞组分主要为自噬前体结构膜、液泡膜、过氧化物酶体等;下调DEGs富集的细胞组分主要为叶绿体、微管、细胞壁等(Fig. 11 B)。上调DEGs富集的分子功能包括转录因子活性、离子结合、泛素蛋白连接酶活性等;下调DEGs富集的分子功能包括微管结合、过氧化物酶活性、水解酶活性等(Fig. 11 C)。KEGG分析显示,DEGs主要参与角质、栓质和蜡质生物合成、苯丙烷类生物合成、淀粉和蔗糖代谢、光合作用、MAPK信号通路-植物、植物激素信号转导等通路(Fig. 11 D)。
图11. 野生型(WT)和WT(干旱处理)植株间基因表达的全局分析。A. WT(干旱处理)-vs-WT组中所有注释的差异表达基因(DEGs)的基因本体论(GO)富集的生物学过程分析。B. WT(干旱处理)-vs-WT组中所有注释DEGs的GO富集的细胞组分分析。C. WT(干旱处理)-vs-WT组中所有注释DEGs的GO富集的分子功能分析。WT(干旱处理)-vs-WT,干旱处理后的野生型植株对野生型植株。根据p值选择了20个最显著的GO类别。D. WT(干旱处理)-vs-WT组中注释DEGs的京都基因与基因组百科全书(KEGG)分析。WT(干旱处理)-vs-WT,干旱处理后的野生型植株对野生型植株。根据q值选择了20个最显著的KEGG通路。
3.11 TG(干旱处理)-vs-TG组差异表达基因的GO和KEGG分析
上调GO富集:富集于非生物刺激响应、胁迫响应、过氧化氢响应等。
下调GO富集:富集于氧化脂生物合成过程、碳水化合物代谢过程、光合作用等。
KEGG通路富集:主要富集于角质、栓质和蜡质生物合成、光合作用、MAPK信号通路-植物等。
基于GO分析,上调的DEGs主要富集的生物学过程包括对非生物刺激的响应、对胁迫的响应、对过氧化氢的响应、对活性氧的响应等;下调的DEGs主要富集的生物学过程包括氧化脂生物合成过程、碳水化合物代谢过程、细胞壁组织、光合作用等(Fig. 12 A)。上调DEGs富集的细胞组分主要为叶绿体类囊体膜、光系统I、光系统II等;下调DEGs富集的细胞组分主要为叶绿体、细胞壁、质外体等(Fig. 12 B)。上调DEGs富集的分子功能包括转录因子活性、氧化还原酶活性、铁离子结合等;下调DEGs富集的分子功能包括过氧化物酶活性、纤维素合酶活性、水解酶活性等(Fig. 12 C)。KEGG分析显示,DEGs主要参与角质、栓质和蜡质生物合成、光合作用、MAPK信号通路-植物、类黄酮生物合成、淀粉和蔗糖代谢、植物激素信号转导等通路(Fig. 12 D)。
图12. 转基因植株(TG)和TG(干旱处理)植株间基因表达的全局分析。A. TG(干旱处理)-vs-TG组中所有注释的差异表达基因(DEGs)的基因本体论(GO)富集的生物学过程分析。B. TG(干旱处理)-vs-TG组中所有注释DEGs的GO富集的细胞组分分析。C. TG(干旱处理)-vs-TG组中所有注释DEGs的GO富集的分子功能分析。TG(干旱处理)-vs-TG,干旱处理后的转基因植株对转基因植株。根据p值选择了20个最显著的GO类别。D. TG(干旱处理)-vs-TG组中注释DEGs的京都基因与基因组百科全书(KEGG)分析。TG(干旱处理)-vs-TG,干旱处理后的转基因植株对转基因植株。根据q值选择了20个最显著的KEGG通路。
3.12 生长发育及非生物胁迫相关差异表达基因的分析
在TG植株中观察到的表型促使研究聚焦于与生长和胁迫响应相关的DEGs,以阐明EpMYB影响植物发育和抗逆性的分子机制。在TG-vs-WT比较组中鉴定出一些与生长、发育和干旱胁迫响应相关的DEGs(Table 2)。此外,通过比较TG(干旱处理)-vs-TG组与WT(干旱处理)-vs-WT组,还鉴定出一些表达水平差异显著的基因,这些基因可能与转基因匍匐翦股颖抗旱性的提高密切相关(Table 3)。
首次实现了跨界基因的功能应用:本研究是第一次将一个真菌源的MYB基因(EpMYB,来源于石果衣)成功应用于高等植物(匍匐翦股颖)中,并证实其能够显著提升植物的非生物胁迫(干旱)耐受性。这突破了传统上主要利用植物内源基因进行遗传改良的局限,开创了跨生物界(真菌-植物)利用基因资源改良作物性状的先例。
提出并验证了全新的抗逆基因挖掘策略:研究不再局限于从模式植物或近缘物种中寻找抗逆基因,而是将目光投向了能够在极端干旱环境中生存的耐逆微生物(石果衣)。通过从这类生物中筛选功能基因并应用于植物,本研究提出并成功验证了一种全新的、从极端环境微生物基因库中挖掘宝贵基因资源的创新策略,为未来植物抗逆育种开辟了崭新的基因来源。
揭示了外源真菌转录因子在植物中的多效性调控网络:研究不仅证明了EpMYB基因的有效性,更深入地揭示了其在植物体内的复杂作用机制。结果表明,这单个真菌转录因子能够作为一个“总开关”,系统性地、多途径地调控宿主植物的生长发育和胁迫响应。它能够系统性地整合并调控宿主植物内源的多个关键生命过程,其影响范围涵盖了从激素信号平衡、木质素与蜡质等物理屏障的构建,到光合作用效率、抗氧化防御及渗透调节等核心生理代谢途径,为理解基因功能的网络调控提供了新的视角。
EpMYB基因对匍匐翦股颖的生长速率、叶片数量、节间长度和最大叶片长度有显著促进作用,但部分叶片出现严重扭曲现象。
转基因植物的抗氧化酶含量、木质素含量和抗旱能力显著增强。
RNA-seq分析显示,EpMYB基因通过调控光合作用、蜡质生物合成、脂质代谢和类黄酮生物合成等途径增强抗旱能力。
EpMYB基因显著上调了与植物生长、发育和抗逆性相关的差异表达基因(DEGs),如木质素合成基因HCT4、蜡质合成基因CER1和脂质代谢相关基因GPAT。
EpMYB基因通过调控植物激素(如IAA、BR和ZR)的水平影响植物生长和应对非生物胁迫的能力。
研究验证了从耐旱微生物中选择特定功能基因并应用于植物以增强抗逆性的方法的可行性。
EpMYB基因显著促进了转基因植物的光合作用,特别是通过调控ERECTA基因和ROC8基因影响植物形态发生。
EpMYB基因通过调控脂质合成和代谢、蜡质合成以及不饱和脂肪酸代谢显著增强了植物的抗旱能力。
实验室条件下的研究结果需要通过田间试验验证其在自然环境下的表现。
该研究提供了关于EpMYB基因在植物抗旱性改良中的理论基础和技术资源,同时提出了基于耐旱微生物基因的跨学科创新方法。