闫丹丽1,武俊瑞1,刘巨龙2,李欣雨1,王雨欣1,李彤1,杨海鑫1,李辉1,马溆雯1,王晨元2,乌日娜1*
(1.沈阳农业大学食品学院 辽宁省食品发酵技术工程研究中心 沈阳市微生物发酵技术创新重点实验室,沈阳 110866;2.蒙牛高科乳制品(北京)有限责任公司,北京 102200)
摘 要:肠道炎症性疾病以肠道屏障损伤、菌群失调及免疫稳态失衡为核心病理特征,其中巨噬细胞M1/M2极化失衡是驱动炎症发生发展与慢性化的关键环节。益生菌胞外囊泡(Probiotic-derived extracellular vesicles,PEVs)作为益生菌分泌的纳米级囊泡,是一类新型后生元,可携带性成分跨越肠道屏障靶向巨噬细胞,在调控肠道炎症中展现出独特潜力。本文系统阐述PEVs通过调控巨噬细胞免疫代谢以诱导其M2型极化,进而缓解肠道炎症这一核心机制的研究进展。重点介绍了PEVs的释放机制与化学组成、关键效应分子通过模式识别受体靶向巨噬细胞的途径及其调控关键信号通路与代谢网络的分子机制。文章为深入理解PEVs调控肠道免疫稳态的分子机制提供了新视角,也为开发基于PEVs的功能性乳品及肠道炎症干预策略提供理论参考。
关键词:益生菌;胞外囊泡;后生元;巨噬细胞极化;肠道炎症
中图分类号:TS201.3
文献标识码:A
DOI:10.19827/j.issn1001-2230.2026.05.001
基金项目:国家自然科学基金项目(32172279);蒙牛乳业基础研究项目(202509046052024095);辽宁省“兴辽英才计划”项目(XLYC2402005;XLYC2213026);辽宁省科学技术计划项目(2024JH2/101900005);沈阳农业大学引进人才(高层次)科研启动费项目(2023YJRC002);沈阳市科技创新平台项目(21-103-0-14、21-104-0-28)。 作者简介:闫丹丽(1996年—),女,博士研究生,研究方向为食品营养与健康。通信作者:乌日娜,女,教授,博士生导师,研究方向为食品营养与健康。闫丹丽,武俊瑞,刘巨龙,等.益生菌胞外囊泡调节巨噬细胞极化缓解肠道炎症研究[J].中国乳品工业,2026,54(5):5-12,19.DOI:10.19827/j.issn1001- 2230.2026.05.001.
YAN Danli,WU Junrui,LIU Julong,et al Research progress on probiotic-derived extracellular vesicles modulating macrophage polarization to alleviate intestinal inflammation[J].China Dairy Industry,2026,54(5):5-12,19.DOI:10.19827/j.issn1001-2230.2026.05.001.
0 引 言
肠道炎症是肠道屏障、菌群和免疫三者构成的肠道稳态失衡后引起的病理反应,易引发炎症性肠病(Inflammatory bowel disease,IBD)、肠易激综合征(Irritable bowel syndrome,IBS)等慢性肠道疾病。巨噬细胞作为肠道固有免疫的核心细胞,在识别病原体或损伤信号后激活NF-κB、MAPK通路,大量分泌TNF-α、IL-6等促炎因子,形成炎症恶性循环。临床研究发现,IBD患者肠道黏膜中,M1型巨噬细胞过度极化,释放的活性氧、一氧化氮直接损伤上皮细胞DNA,导致黏膜糜烂和溃疡形成[1]。因此,调控巨噬细胞极化方向已成为缓解肠道炎症的重要策略。
益生菌的免疫调控功能已获广泛证实。植物乳植杆菌(Lactiplantibacillus plantarum)、双歧杆菌(Bifidobacterium)等益生菌可通过抑制致病菌增殖,促进有益菌生长、提高紧密连接蛋白ZO-1、Occludin表达水平,增强黏液层厚度,修复肠道屏障[2]。然而,传统益生菌依赖活菌定植,口服后90%以上被胃酸和胆汁灭活,且菌株特异性强、抗炎效果差异显著,这些局限性促使研究者将目光转向益生菌来源的后生元。益生菌胞外囊泡(Probiotic-derived extracellular vesicles,PEVs)是益生菌主动释放的纳米级脂质双层囊泡,可穿越上皮屏障,将蛋白质、核酸、脂质等活性成分高效递送至巨噬细胞,具有免疫原性低、稳定性高的特点[3]。研究表明,PEVs可通过调节巨噬细胞极化从而调节肠道炎症。黏膜乳杆菌EVs可通过诱导巨噬细胞从M1型极化转向M2型极化,显著降低仔猪肠道炎性损伤评分[4]。罗伊氏乳杆菌膜囊泡能促进嗜黏蛋白阿克曼菌(Akkermansia muciniphila)增殖、激活HIF-1信号通路、上调紧密连接蛋白表达,维持肠道免疫稳态[5]。这些研究为靶向巨噬细胞极化以干预肠道炎症提供了基于微生物源纳米囊泡的新策略,更为开发靶向免疫调节功能性发酵乳制品提供了新方向。
鉴于PEVs在调节巨噬细胞免疫代谢、重塑肠道免疫稳态中的独特优势,近年来相关研究取得了重要进展。然而,不同益生菌菌株来源EVs的功能差异、其调控巨噬细胞极化的机制以及作为功能性乳制品配料的应用前景尚缺乏系统梳理。本文围绕PEVs通过调控巨噬细胞免疫代谢诱导M2型极化、进而缓解肠道炎症这一核心机制研究进行综述,重点阐述PEVs携带的关键效应分子通过模式识别受体靶向巨噬细胞的过程,及其调控NF-κB、MAPK-STAT3及代谢重编程等多条信号通路的分子机制,并探讨了PEVs作为新型后生元在功能性乳制品开发中的产业化前景,以期为PEVs在食品工业及肠道健康领域的转化应用提供理论支撑。
1 PEVs的生物学特征
PEVs是益生菌分泌的纳米级脂质双层膜结构,作为介导菌群-宿主互作的新型效应分子,功能多样性源于其独特的来源、形成机制及复杂的化学组成。这些基础生物学特征共同决定了PEVs的稳定性、生物相容性及靶向调控免疫细胞的能力。
1.1 来源与形成机制
PEVs的形成机制与亲本菌的细胞壁结构密切相关,主要根据来源菌的革兰氏染色特性进行分类。革兰氏阴性菌PEVs的形成主要依赖外膜与肽聚糖层的交联松弛。当OmpA、Lpp等交联蛋白功能减弱时,外膜局部膨出并脱落形成囊泡,主要分为3种途径:其一,外膜囊泡(Outer membrane vesicles,OMVs),由外膜直接出芽脱落形成,是最常见的类型;其二,外-内膜囊泡(Outer-inner membrane vesicles,OIMVs),当肽聚糖层局部降解后,内膜突入周质空间并被外膜包裹,形成具有双层膜结构的囊泡;其三,爆炸性外膜囊泡(Explosive outer membrane vesicles,EOMVs),在抗生素或氧化应激等刺激下,噬菌体来源的内溶素被激活并降解细胞壁,导致细菌裂解,膜碎片自发重排形成包裹全部细胞组分的爆炸性囊泡[6]。相比之下,革兰氏阳性菌没有外膜,且其肽聚糖层较厚,形成了一层物理屏障,阻止了EVs释放,使其囊泡形成曾长期被忽视。研究表明,双歧杆菌属及乳杆菌属等多种革兰氏阳性益生菌PEVs主要通过细胞膜出芽并借助肽聚糖水解酶局部降解细胞壁而形成,称为细胞质膜囊泡(Cytoplasmic membrane vesicles,CMVs)[7]。
1.2 化学组成与功能特性
PEVs是一个复杂的活性分子库,其调控巨噬细胞极化、缓解肠道炎症的功能由携带的蛋白质、核酸、脂质及代谢物共同决定。不同菌株来源的PEVs因化学组成差异,呈现出多样化的免疫调节功能。
蛋白质是PEVs的核心功能成分。大肠杆菌Nissle 191EVs中的OmpA、OmpC等结构蛋白可促进与免疫细胞的相互作用[8]。MANDELBAUM等[9]发现长双歧杆菌长亚种(Bifidobacterium longum subsp. longum)AO44分泌的EVs中高度富集的ABC转运蛋白、群体感应蛋白和细胞外溶质结合蛋白,这些组分可能参与其抗炎作用。此外蛋白质组成随细菌生长阶段动态变化,多形拟杆菌EVs中脂蛋白和细胞质蛋白的相对丰度呈时间依赖性,脂化后半胱氨酸C端带负电荷的氨基酸是蛋白质分选进入囊泡的关键[10]。
核酸(DNA、RNA)是PEVs行使跨物种基因调控功能的关键。PEVs中DNA可介导基因水平转移,而小非编码RNA通过调控宿主细胞基因表达诱导表型变化。研究发现,植物乳植杆菌胞外囊泡通过递送高丰度的cbn-let-7靶向抑制脂质代谢酶ACSL4表达,从而阻断巨噬细胞铁死亡[11]。脆弱拟杆菌EVs携带的miR-5119样小RNA通过直接靶向PD-L1 mRNA抑制其表达、进而下调GSDMD介导的性粒细胞胞外诱捕网释放,减轻肠道炎症损伤,促进杯状细胞和肠道干细胞再生[12]。这些发现表明,PEVs膜结构为内部核酸提供卓越保护,使其在胃肠道环境中保持高度稳定,荧光标记RNA研究显示,EVs递送的细菌小RNA可在宿主细胞内维持12~24 h,足以调控靶基因表达。
脂质不仅构成PEVs的膜骨架,其本身也是重要信号分子。脂质组学分析显示,PEVs携带多种磷脂,包括磷脂酸、磷脂酰胆碱、磷脂酰乙醇胺和磷脂酰甘油,这些成分通过维持膜结构流动性和稳定性,保障PEVs的功能完整。不同菌群来源PEVs的脂质组成具有显著差异。PEVs的脂质组成与其外膜高度相似,主要由甘油磷脂、心磷脂和脂多糖构成,这些脂质通过维持脂质双层的流动性和稳定性保障囊泡功能。革兰氏阳性细菌的CMVs富含质膜脂质,如鼠李糖乳酪杆菌(Lacticaseibacillus rhamnosus)的CMVs含有磷脂酰胆碱,该成分可与宿主肠上皮细胞脂质筏融合,增强抗炎性miRNA的递送效率[13]。不脂质组成显著影响PEVs与宿主细胞的融合效率,大肠杆菌OMVs中的磷脂不对称性增强其与上皮细胞的结合能力,而VacJ/Yrb系统缺陷导致的脂质不对称性破坏则会降低内容物递送效率[14]。
多糖与小分子代谢物同样参与PEVs免疫调控。大肠杆菌(Escherichia coli)O157:H7外膜囊泡所含LPS可与宿主肠上皮细胞表面的CD14受体结合,诱导细胞焦亡、破坏肠道屏障功能并干扰宿主细胞脂质合成[15]。肽聚糖是革兰氏阳性菌与阴性菌PEVs的共同组分。大肠杆菌OMVs中的肽聚糖片段通过NOD1激活宿主肠上皮细胞免疫应答,促进宿主细胞分泌抗菌肽,从而调控肠道菌群组成与代谢[19]。脆弱拟杆菌(Bacteroides fragilis)OMVs中的荚膜多糖A通过TLR2激活宿主树突状细胞,诱导调节性T细胞分化及IL-10分泌,同时调节宿主肠道葡萄糖代谢[16]。除多糖外,PEVs还可转运吲哚、短链脂肪酸、乳酸等小分子代谢物。普拉梭菌(Faecalibacterium prausnitzii)EVs含有肌苷和琥珀酸,卷曲乳杆菌(Lactobacillus crispatus)EVs携带的D-乳酸可促进M2型巨噬细胞极化和伤口愈合[17]。这些小分子代谢物可直接被宿主细胞利用或触发信号转导,发挥抗炎、促进上皮修复等作用。
1.3 稳定性与生物相容性
优异的稳定性和生物相容性是PEVs作为潜在后生元制剂应用于功能性乳制品开发的重要前提。研究表明,PEVs能较好耐受模拟胃肠道的恶劣环境(低pH、胆盐、消化酶),在4 ℃或-80 ℃下可长期稳定保存,甚至能耐受一定热处理,这为其在乳品加工中的应用提供了可能。同时,PEVs对肠上皮细胞和免疫细胞表现出低细胞毒性,具有良好的生物安全性[18]。这些特性使PEVs能够穿越肠道屏障,将携带的活性分子高效递送至以巨噬细胞为代表的靶细胞,为靶向调控肠道免疫稳态奠定了基础。
2 巨噬细胞极化与肠道炎症
2.1 巨噬细胞具有可塑性
巨噬细胞是免疫系统中可塑性最强的细胞群体,这种可塑性使其能感知微环境信号并动态调整表型与功能,从而在维持组织稳态、启动炎症反应及促进组织修复中发挥核心作用。当识别病原体相关分子模式(Pathogen-associated molecular patterns,PAMPs)或损伤相关分子模式(Damage-associated molecular pattern,DAMPs)时,巨噬细胞通过模式识别受体(Pattern recognition receptors, PRRs)激活胞内免疫信号通路,产生以IL-6和TNF-α为代表的促炎因子及趋化因子。这些因子招募中性粒细胞、NK细胞及T淋巴细胞,并通过IFN-γ的分泌进一步增强促炎反应,形成正反馈放大效应,从而增强其杀菌、抗病毒和抗肿瘤活性[19]。而在组织修复微环境中,巨噬细胞则产生IL-4、IL-10、IL-13和TGF-β等抗炎细胞因子,招募Th2细胞、嗜酸性粒细胞和嗜碱性粒细胞,进而刺激巨噬细胞分泌IL-10,形成抗炎正反馈调节。这种根据环境信号改变功能表型的过程称为极化[20]。
2.2 巨噬细胞极化失衡调节肠道炎症
巨噬细胞极化的动态平衡,即促炎M1型与抗炎M2型的分化调控,是决定肠道炎症进程与转归的核心环节。M1型巨噬细胞通常由IFN-γ、TNF-α或LPS等信号诱导,通过激活NF-κB和JAK/STAT1通路,大量分泌IL-1β、IL-6、TNF-α等促炎因子以及一氧化氮和活性氧。这一表型在清除病原体中发挥关键作用,但过度活化会严重损伤肠道上皮完整性,增加肠道通透性,并招募更多炎症细胞浸润,形成促炎恶性循环[21]。临床与实验数据均证实,在IBD患者和DSS结肠炎小鼠模型中,M1型巨噬细胞比例及促炎因子水平均与炎症严重程度呈正相关。与之相对,M2 型巨噬细胞在IL-4、IL-13或IL-10等信号诱导下活化,通过分泌IL-10、TGF-β及表达精氨酸酶-1,发挥抗炎、促进上皮增殖与组织重塑的作用,是炎症恢复期黏膜修复的关键。因此,巨噬细胞极化平衡的打破是肠道炎症的核心机制,将极化方向向M2型转向已成为重要的治疗策略,这为PEVs作为天然靶向调控工具的应用奠定了理论基础。
3 PEVs在肠道微生态调控中的核心功能
益生菌胞外囊泡作为益生菌功能的重要载体,通过调节肠道菌群平衡、增强肠道屏障完整性及调控肠道免疫应答这三大核心功能,协同维持肠道微生态稳态,三者相互关联、相互促进,共同构成了PEVs维持肠道健康的综合调控网络。
3.1 调节肠道菌群平衡
肠道菌群失衡是诱发肠道炎症的关键因素。PEVs通过抑制致病菌与促进有益菌的双向调控策略,有效恢复菌群稳态,为缓解肠道炎症奠定微生态基础。
肠道菌群失衡是诱发肠道炎症的关键因素。PEVs通过抑制致病菌与促进有益菌的双向调控策略,有效恢复菌群稳态,为缓解肠道炎症奠定微生态基础。在抑制致病菌方面,PEVs一方面发挥直接抗菌作用,如大肠杆菌Nissle 1917来源的OMVs可分泌抗菌物质直接破坏肠球菌细胞膜,使其存活率降低60%[23]。另一方面干扰致病菌群体感应系统,植物乳植杆菌来源的EVs通过抑制luxS基因表达和自诱导物-2的产生,下调单增李斯特菌(Listeria monocytogenes)的毒力与黏附相关基因,从而削弱其对肠上皮的侵袭能力[24]。在促进有益菌方面,PEVs通过营养支持、代谢激活和环境优化,选择性富集阿利斯蒂普斯属(Alistipes)等有益菌,抑制变形菌门(Proteobacteria)等促炎菌群,同时恢复短链脂肪酸水平并优化肠道pH环境。研究表明,PEVs通过调节菌群组成与代谢功能缓解肠道炎症。植物乳植杆菌Q7来源的EVs可降低变形菌门丰度,增加双歧杆菌属和毛球菌科(Muribaculaceae)丰度[25]。普拉梭菌来源的OMVs可增加磷脂酰胆碱等关键代谢物水平,减轻病毒感染所致肠道损伤[26]。
3.2 强化肠道屏障完整性
肠道屏障完整性是防止有害物质易位的结构基础。PEVs可协同强化其物理与生物屏障功能。
3.2.1 强化物理屏障
肠道物理屏障的核心是肠上皮细胞及其间的紧密连接蛋白。PEVs可通过调控紧密连接蛋白表达与组装、平衡上皮细胞增殖与凋亡两大途径增强其功能。
在调控紧密连接蛋白方面,不同来源的PEVs通过激活特定的细胞内信号通路实现这一目标。如发酵粘液乳杆菌(Limosilactobacillus fermentum)SLAM 216来源EVs处理小鼠小肠类器官后,可显著上调紧密连接相关基因Claudin、Occludin、ZO-1的表达,促进肠道发育与成熟[27]。此外,植物乳植杆菌HY7714来源EVs可通过调控MyD88/mTOR/NF-κB信号通路,修复受损的肠道细胞结构并恢复紧密连接完整性,同时下调炎症细胞因子表达[28]。大肠杆菌Nissle 1917来源EVs通过激活TLR2信号通路上调修复因子TFF3表达,增强紧密连接蛋白亚细胞定位并刺激Caco-2细胞的伤口愈合疫[29]。
在调节上皮细胞增殖与凋亡平衡方面,PEVs携带的多种活性分子可加速损伤修复。食淀粉乳杆菌(Lactobacillus amylovorus)QC1H来源EVs激活肠道AHR/IL-22信号通路,抑制促炎因子产生、降低肠道通透性、增加紧密连接蛋白表达显著缓解AFB1诱导的肠道损伤[13]。此外,PEVs携带的EGF样蛋白可促进细胞增殖,而特定miRNA则可抑制细胞凋亡,二者共同促进黏膜修复[30]。
3.2.2 加固生物屏障
肠道生物屏障是由肠道上皮分泌的黏液层、抗菌物质与共生菌群共同构成的动态防御体系,PEVs主要通过促进黏液分泌和诱导抗菌物质加固这一屏障。在促进黏液分泌方面,多项研究证实PEVs可显著上调黏蛋白MUC2表达。如嗜粘蛋白阿克曼氏菌来源EVs干预可显著升高结肠炎小鼠肠道MUC2表达水平,改善黏液完整性,降低肠道通透性[31]。在诱导抗菌物质方面,罗伊氏粘液乳杆菌(Limosilactobacillus reuteri)来源EVs携带可作为TLR2激动剂的脂磷壁酸,可调节宿主免疫应答,其携带的5’-核苷酸酶可将AMP转化为信号分子腺苷,发挥免疫调节作用[32]。综上,PEVs通过促进黏液分泌及诱导抗菌物质等多重机制,协同加固肠道生物屏障,为维持肠道稳态奠定基础。
3.3 调控肠道免疫应答
巨噬细胞作为肠道固有层中数量最丰富的免疫细胞,是维持肠道稳态的核心效应细胞;而树突细胞则作为连接先天免疫与适应性免疫的关键桥梁,其功能状态直接影响后续T细胞应答方向。PEVs对这2类细胞的精准调控构成了其先天性免疫调节的基础。研究表明,PEVs能抑制巨噬细胞的过度活化,减少促炎因子释放,并促进其向具有抗炎和组织修复功能的M2型极化。这一调控具有菌株特异性,不同来源PEVs通过携带不同效应分子发挥作用。如副干酪乳酪杆菌(Lacticaseibacillus paracasei)、嗜酸乳杆菌(Lactobacillus acidophilus)、德氏乳杆菌保加利亚亚种(Lactobacillus delbrueckii subsp. bulgaricus)、瑞士乳杆菌(Lactobacillus helveticus)、植物乳植杆菌等部分乳酸菌来源PEVs可激活TLR2,并与FPR2协同作用,通过抑制NF-κB和MAPKs通路、调控STAT3磷酸化及钙离子信号,显著增加IL-10分泌,协同诱导M2型极化[33]。对于树突状细胞,约氏乳杆菌(Lactobacillus johnsonii)N6.2来源的磷脂可诱导树突状细胞维持未成熟样表型,降低共刺激分子和MHC分子的表面表达从而发挥免疫抑制功能[34]。
在适应性免疫调节方面,PEVs通过调控树突状细胞功能间接影响T细胞应答。PEVs诱导的耐受型树突状细胞与T细胞相互作用时,可抑制Th1、Th17等促炎亚型的效应功能,同时诱导T细胞无反应性相关基因表达,促进免疫耐受形成。进一步研究发现,约氏乳杆菌来源EVs可显著减少DSS结肠炎小鼠结肠中Th17细胞比例、增加Treg细胞数量,恢复Th17/Treg平衡,从而缓解肠道炎症[35]。此外,PEVs还能增强黏膜免疫,通过激活B细胞促进分泌型免疫球蛋白A的产生,从而在黏膜表面建立一道重要的免疫防线[36]。
4 PEVs调节巨噬细胞极化缓解肠道炎症机制
4.1 PEVs穿越生物屏障靶向巨噬细胞途径
PEVs 需依次突破黏液层、肠上皮屏障2道肠道生物屏障,再通过特异性靶向与高效摄取机制富集于巨噬细胞,其高效性源于纳米尺度和表面分子协同作用。
穿透黏液层是PEVs面临的第一道关卡。肠道黏液层由高糖基化黏蛋白构成网状结构,可有效截留病原体和颗粒物。PEVs凭借其纳米尺寸(通常50~300 nm)和磷脂双分子层结构,可通过被动扩散有效穿透黏液网络,其效率与粒径成反比。干酪乳酪杆菌(Lacticaseibacillus casei)BL23来源EVs能以剂量依赖性方式与黏蛋白结合,这为其穿越黏液层提供了分子基础。部分PEVs还能分泌黏液溶解酶适度降解黏液,进一步提高渗透效率[37]。研究表明,细菌EVs的黏液穿透能力显著优于许多游离药物和合成纳米颗粒。如与被动穿透的合成纳米颗粒不同,嗜粘蛋白阿克曼氏菌EVs作为口服siRNA的有效工具通过降解肠道黏蛋白实现主动黏液穿透,使其穿越肠道屏障能力提升约4.6倍[38]。普拉梭菌PEVs口服后12 h,超过90%的结肠巨噬细胞可摄取该囊泡,且PEVs 在肠道炎症部位的富集程度显著高于健康肠道[39]。
PEVs 通过受体介导的内吞与特异性胞内转运实现跨上皮递送。其表面的 OmpA、黏附素等黏附蛋白与肠上皮细胞的转铁蛋白受体、整合素等结合后,被内化形成网格蛋白包被囊泡。工程化大肠杆菌Nissle 1917来源的外膜囊泡能够通过胞饮作用和动力蛋白依赖途径完成跨上皮运输,进入循环系统[40]。嗜粘蛋白阿克曼氏菌在胞内具有独特的高尔基体-内质网运输通道,能够将内容物精准送抵细胞内的蛋白质合成中心,有效规避传统的内涵体-溶酶体降解途径,从而提高货物递送效率[41]。
进入固有层后,PEVs通过趋化因子梯度和表面配体-PRRs结合双重机制富集于巨噬细胞。一方面,炎症部位高表达的CCL2、CCL5吸引表面携带CCR2、CCR5的PEVs向病灶聚集。另一方面,PEVs表面的脂磷壁酸、脂多糖、肽聚糖片段分别与巨噬细胞的TLR2、TLR4、NOD1/2等PRR特异性结合,同时启动摄取与信号激活ADDIN。体内分布研究证实了PEVs的巨噬细胞靶向性。长双歧杆菌长亚种来源EVs和植物乳植杆菌WCFS1来源EVs经过静脉注射后,主要被肝脏F4/80+巨噬细胞和脾脏CD169+巨巨噬细胞高效捕获[42]。此外,工程化改造可增强靶向性。通过基因工程修饰PEVs表面分子可进一步提高其巨噬细胞靶向效率,例如表达抗CD206纳米抗体的PEVs能特异性识别巨噬细胞高表达的甘露糖受体,显著提高摄取效率和功能调控特异性[43]。
巨噬细胞对PEVs的摄取具有尺寸依赖性:大于1 μm的通过吞噬作用摄取,0.5~1.0 μm的通过巨胞饮作用摄取,小于0.5 μm的通过网格蛋白介导的内吞摄取。其中网格蛋白介导的内吞是主要途径且具有保守性,长双歧杆菌长亚种、丁酸梭菌(Clostridium butyricum)和植物乳植杆菌 WCFS1 等不同来源 PEVs 在RAW264.7巨噬细胞中均以此途径为主[44]。双歧杆菌PEVs的内化过程则需动力蛋白参与胞内转运以实现货物释放[45]。巨胞饮作用是PEVs摄取的次要途径,尤其当PEVs浓度较高或尺寸为100~400 nm 时发挥重要作用。大肠杆菌Nissle 1917来源OMVs可诱导巨噬细胞细胞膜皱褶形成杯状结构,将大量囊泡包裹进入胞内形成巨胞饮体,该过程依赖Rac1和Pak1信号分子的激活,且与囊泡表面脂多糖含量正相关[46]。小窝蛋白介导的内吞主要针对20~50 nm的小型PEVs。鼠李糖乳酪杆菌 PEVs 可与巨噬细胞膜小窝蛋白-1结合完成内化,该途径能耗低、对膜结构破坏小,能保护货物生物活性,小窝蛋白-1敲除后巨噬细胞对其摄取效率及抗炎效应均显著下降[47]。膜融合途径是部分PEVs递送货物的独特方式。罗伊氏粘液乳杆菌PEVs的膜脂质组成与巨噬细胞膜高度相容,其表面的磷脂酰丝氨酸可与巨噬细胞表面的Annexin A5结合,促进膜融合并将3-羟基丙醛直接递送至巨噬细胞线粒体,避免内体酸化对货物分子的降解[48]。这些摄取机制并非相互排斥,同一PEVs群体可通过多种途径进入巨噬细胞且被摄取后,其货物分子通过内体逃逸、膜融合或内体破裂等方式释放到巨噬细胞胞质中,进而启动极化调控信号。这一过程的效率直接影响其抗炎效应发挥。
4.2 PEVs调节巨噬细胞极化机制
PEVs被巨噬细胞摄取后,其携带的多种活性成分通过干预多条关键信号通路,协同促使巨噬细胞向M2型极化。这些通路主要包括PPRs介导的信号通路和免疫代谢重编程2个层面,二者协同抑制M1型极化、促进M2型转换,共同构成PEVs调控巨噬细胞极化分子网络。
4.2.1 PPRs介导的信号通路
PRR与PEVs表面配体特异性结合是极化调控起始环节,不同来源PEVs因表面分子组成差异,可激活差异化PRR信号通路,表现出显著菌株特异性。
TLR2是介导PEVs免疫调控的核心受体之一。鼠李糖乳酪杆菌PEVs通过激活TLR2信号通路,招募MyD88适配器分子,激活下游IRAK-TRAF6信号级联,促进IL-10等抗炎因子分泌,诱导巨噬细胞向M2型极化[49]。植物乳值杆菌EVs中脂蛋白来源N-酰基氨基也可激活TLR2,调节IL-6、IL-12、IL-10等细胞因子分泌水平,进而刺激未致敏T细胞向辅助T细胞分化并诱导B淋巴细胞产生IgA[50]。戊糖片球菌以TLR2依赖性方式促进巨噬细胞M2极化和髓源抑制细胞分化上调IL-10、Arg1和PD-L1的表达,同时抑制活化的T细胞增殖,防止DSS诱导的结肠炎小鼠结肠长度缩短,并保护其隐窝结构完整性[51]。
TLR4信号通路在PEVs调控中同样发挥重要作用。加氏乳杆菌(Lactobacillus gasseri)来源EVs的膜表面脂多糖与巨噬细胞TLR4结合后,通过抑制TLR4/NF-κB信号通路的激活,显著降低线粒体超氧化物和活性氧水平,抑制NO、PGE2及其关键酶iNOS和COX-2的表达,同时下调IL-6、TNF-α和IL-1β等促炎因子分泌,从而实现从M1向M2表型转换[52]。
胞内受体信号通路也参与PEVs免疫调控。大肠杆菌来源OMVs可被巨噬细胞胞内NOD1受体识别,通过激活下游RIP2激酶,调控NF-κB和MAPK通路,增强肠上皮细胞紧密连接蛋白表达,从而减轻炎症因子对肠黏膜的损伤[53]。约氏乳酸杆菌-NII菌株来源EVs通过抑制ERK1/2的磷酸化,阻断NLRP3炎症小体组装和激活,减少IL-1β成熟和释放,从而缓解肠毒素性大肠杆菌K88引起的巨噬细胞功能障碍和肠道上皮屏障损伤[54]。植物乳植杆菌胞外囊泡可携带色氨酸激活AhR受体,AhR受体被激活后能促进IL-22产生,从而帮助维持肠道屏障功能、调节肠道蠕动和内脏敏感性,缓解肠易激综合征症状[55]。
除PRRs外,PEVs还可通过其他分子调控信号通路。约氏乳杆菌EVs携带的GAPDH可抑制p38 MAPK磷酸化,解除对STAT3的抑制性调控,促进STAT3活化并启动IL-10、Arg-1等M2标志基因转录[56]。乳酸乳球菌来源的EVs携带miR-146b靶向激活STAT6/PPARγ信号通路,抑制NF-κB活性,进而诱导巨噬细胞从M1表型向M2表型转换[57]。丁酸梭菌(Clostridium butyricum)来源EVs携带miR-199a-3p接靶向结合MAPK4,阻断MAPK和NF-κB信号级联反应,减少促炎细胞因子产生,减轻DSS诱导的结肠炎症[58]。淀粉芽胞乳杆菌(Sporolactobacillus amyloliticus)来源EVs通过上调NF-κB抑制蛋白IκBα的表达,有效抑制NF-κB核转位和转录活性,从而促进免疫抑制性M2巨噬细胞表型形成[59]。这些研究表明,PEVs 通过精准激活巨噬细胞信号通路,实现极化平衡调控,为肠道炎症干预提供了靶向策略。
4.2.2 免疫代谢重编程
巨噬细胞极化伴随代谢重构:M1型依赖有氧糖酵解,而M2型偏好氧化磷酸化和脂肪酸氧化。PEVs能通过调节代谢关键酶和通路,促使巨噬细胞代谢模式向M2型转换。PEVs 通过调控巨噬细胞的糖酵解、氧化磷酸化、脂肪酸氧化等核心代谢通路,实现极化表型的重编程。
M1型巨噬细胞的有氧糖酵解是其促炎关键。普拉梭菌EVs可抑制巨噬细胞线粒体功能,降低氧化磷酸化与糖酵解活性,下调PPARγ和ABC转运蛋白表达,减少胆固醇外流,促进抗纤维化M2b亚型极化[60]。同时,EVs被巨噬细胞摄取后,通过上调BRCA1和BRCA2等关键蛋白表达,减少DNA损伤并抑制cGAS-STING炎症通路,从而降低促炎因子产生并增强肠道屏障完整性。大肠杆菌来源EVs被RAW264.7巨噬细胞通过吞噬作用摄取后,激活HIF-1α、mTORC1和NF-κB信号通路,促进代谢从氧化磷酸化向有氧糖酵解转换,同时抑制TCA循环。导致细胞内活性氧、ATP和一氧化氮水平显著升高,最终促进巨噬细胞增殖、迁移和M1极化,增强吞噬杀菌功能[61]。对于M2型巨噬细胞,PEVs通过增强氧化磷酸化和脂肪酸氧化,强化其抗炎修复功能。粘液罗伊氏乳杆菌来源PEVs可递送3-羟基丙醛至巨噬细胞线粒体,抑制线粒体通透性转换孔开放,稳定线粒体膜电位,增强氧化磷酸化活性,促进M2型极化并加速肠道黏膜修复[48]。植物乳植杆菌PEVs则通过激活PPAR γ信号,上调脂肪酸氧化关键酶CPT1A表达,促进脂肪酸氧化,强化M2型巨噬细胞抗炎效应[62]。PEVs还可通过激活核受体NR1H,上调胆固醇外流转运蛋白ABCA1的表达,增强巨噬细胞脂质外流,减少泡沫细胞形成,抑制促炎因子产生,推动M2极化[63]。乳酸杆菌来源EVs通过下调JAK1/STAT1信号轴,增强谷胱甘肽生物合成途径,促进代谢流向线粒体脂肪酸氧化转变,从而支持巨噬细胞维持持久的M2型抗炎表型[64]。脆弱拟杆菌EVs中富含的鞘脂,被细胞摄取后与胞内载脂蛋白L9相互作用,激活甲羟戊酸通路,上调HMG-CoA还原酶活性,通过调节脂质代谢促进抗炎细胞因子IL-10大量分泌,发挥免疫调节作用[65]。
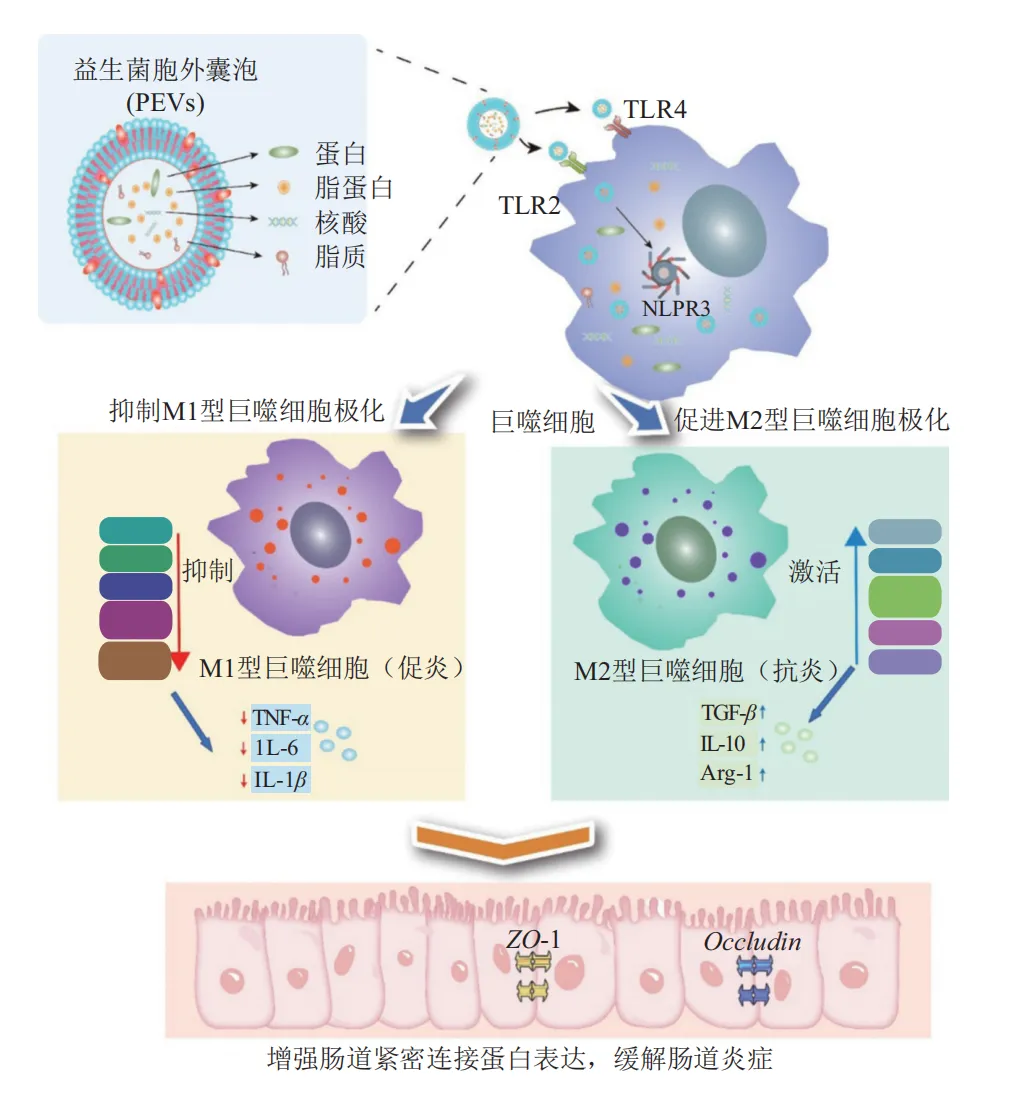
上述研究表明,不同来源PEVs通过精准靶向巨噬细胞糖酵解、氧化磷酸化、脂肪酸氧化及胆固醇代谢等多个核心代谢通路,同时调控HIF-1α、PPARγ、NR1H3等关键信号节点,实现对巨噬细胞极化的精准调控,进而发挥缓解肠道炎症、保护肠道屏障作用,具体分子调控机制见图1。
图1 益生菌胞外囊泡(PEVs)调节巨噬细胞极化缓解肠道炎症分子机制
5 结果与展望
肠道炎症的发生发展与肠道微生态失衡、免疫炎症反应失控密切相关,而PEVs作为新型后生元,凭借独特的磷脂双分子层结构与多活性成分协同优势,成为肠道炎症干预的潜力靶点。PEVs借助纳米尺度和表面分子协同作用,可穿越肠道黏液层和上皮屏障,通过PPRs进入巨噬细胞,其携带的蛋白质、核酸、脂质及代谢物等效应分子,能通过调控 NF-κB、MAPK-STAT3 及代谢重编程等多条通路,协同诱导巨噬细胞从促炎M1型向抗炎M2型极化,进而增强肠道屏障功能、缓解肠道炎症。
在乳制品加工领域,PEVs 展现出区别于传统活菌制剂的独特优势,为功能性乳品开发开辟了新方向。其一,PEVs 具有更优的加工稳定性和货架期适应性。与活菌制剂对高温、酸碱、渗透压变化及防腐剂敏感不同,PEVs 作为无生命的囊泡结构,可抵御胃酸腐蚀和肠道消化酶降解,受热后不易失活,能在保持结构完整的同时顺利递送活性货物;且其在冷藏和常温条件下均不易降解,可长期保留生物活性。这使得含 PEVs 的功能乳品适用范围更广,能够突破冷链限制拓展至常温货架产品,同时确保产品中功能组分的足量与标准化。其二,PEVs 具备更高的安全性。传统活菌制剂需依赖菌株存活发挥作用,若菌株在肠道内过度定植可能打破菌群平衡,对免疫功能低下人群存在条件性感染风险,且可能携带耐药基因转移隐患;而PEVs 不具备增殖能力,仅通过递送生物活性货物发挥作用,无需担心菌株定植引发的微生态紊乱,完全不存在致病性、易位或代谢污染风险,可安全应用于临床营养、特殊医学用途配方食品等对安全性要求极高的领域。其三,PEVs 在适口性上更具优势。传统活菌制剂常带有菌株特有的轻微臭味,可能影响乳制品风味;而PEVs 无味无臭,添加后不改变乳制品原有口感与风味,可广泛适配发酵乳、乳饮料等各类乳制品。此外,PEVs 还能介导跨器官远程调控,且可通过调控发酵条件或基因工程手段定向生产富含特定功能分子的囊泡,实现乳品功能特性的精准定制,满足不同人群的健康需求。
尽管PEVs在肠道炎症干预及功能性乳品开发中展现出显著优势,但目前仍面临规模化生产与纯化技术不成熟、产品加工和储存过程中稳定性不足、标准化制备缺失、机制解析不深入、临床转化滞后等多重挑战。未来,通过多学科交叉合作,建立PEVs的标准化制备与质量控制体系,鉴定核心活性成分并明确其与特定健康功能的对应关系;优化规模化生产纯化工艺及适用于乳品工业的囊泡稳定性保护技术;同时,开展设计严谨的多中心随机对照临床试验,验证含PEVs功能性乳品在肠道炎症干预、免疫调节等场景中的安全性与有效性。通过基础研究、工艺开发和临产验证的协同,有望推动PEVs在功能性乳制品中的广泛应用,为肠道炎症精准预防与治疗提供新型后生元策略,最终实现从基础研究到临床应用的跨越,为肠道健康领域带来新的突破。
(参考文献略)
《中国乳品工业》是由中国轻工业联合会主管,黑龙江省绿色食品科学研究院(原黑龙江省乳品工业技术开发中心)主办的自然科学类学术期刊。《中国乳品工业》期刊1973年创刊,以促进我国乳品学术、技术交流,推动我国乳品工业发展为己任。期刊专注于乳品科学研究及应用技术的报道,注重对行业的前沿领域和交叉学科方面的追踪,对推动该领域的科学研究发挥了重要作用。
入编中文核心期刊,中国科技核心期刊。荣获“第三届国家期刊奖百种重点期刊奖”,“北方优秀期刊称号”,“首届黑龙江省出版奖提名奖”,入选“庆祝中华人民共和国成立七十周年精品期刊”,“中国农业期刊网精品期刊”。被中国知网、万方数据、维普数据、超星数据、长江文库、美国化学文摘(CAS)、美国艾博思科数据库(EBSCO)、英国国际应用生物科学中心数据库(CABI)、英国食品科技文摘(FSTA)、日本科学技术振兴机构数据库(JST)收录。
投稿平台:https://rpgy.cbpt.cnki.net
联系电话:0451-86662740
编辑:王剑飞
马 艳
校对:王 鑫
审核:姜 爽

 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
