作者:特里丰达达博、维托米科利斯、马蒂诺巴锡莱、温琴佐坎迪多
(作者单位:意大利国家研究委员会植物保护研究所,意大利巴里市;巴西利卡塔大学农林环境系统科学系,意大利波坦察市)
关键词:替代防控技术;综合治理;线虫;土壤太阳能消毒;土传病原物;杂草
摘要
施用农药能够高效防控果蔬作物的土传有害生物,但这类药剂对人畜具有毒性,还会在植株与土壤中形成药物残留,加之用药成本偏高,致使农药防控模式既存在安全隐患,经济效益也不理想。与此同时,现行环保法规不断收紧管控要求,多地限制甚至全面禁用大部分土壤处理类农药。在此背景下,非化学防控与减药型植保技术的研发应用日益受到重视。
土壤太阳能消毒是一种无农药防控技术:在高温季节,将塑料薄膜覆盖在湿润土壤表层,依靠密闭增温提升土温,以此灭杀土壤内绝大多数病原真菌、线虫以及杂草种子和幼苗。高温直接灭活有害生物是太阳能消毒发挥杀菌灭害作用的核心机理;除此之外,升温会促使土壤释放有毒挥发性物质,同时驱动土壤微生物群落发生演替,富集大量可拮抗植物病原的有益菌群,进一步强化防控效果。土壤温度与含水量是决定太阳能消毒热力作用效果的关键指标,而地膜材料同样不可或缺:地膜可透过太阳辐射提升地温,同时减少土壤热能的辐射散失与对流损耗。低密度聚乙烯膜、乙烯–醋酸乙烯酯共挤膜的田间控温消毒效果最优,不过市面上多种塑料薄膜材料也均可用于土壤太阳能消毒作业。
土壤经太阳能加温处理后,土壤理化性状得到改良,可溶性养分含量上升,其中可溶性有机质、无机态氮、速效阳离子增幅尤为突出;土壤微生物群落的组成与丰度同步改变,促生菌、病原菌拮抗菌以及根系快速定殖菌群的数量显著提升。得益于上述多重改良作用,大量试验证实,土壤太阳能消毒可连续两个以上种植周期促进作物生长,提升果蔬产量与品质。
众多研究表明,太阳能消毒可强力抑制多数重要植物病原真菌,在多种栽培条件下,能近乎彻底铲除镰刀菌、疫霉菌、腐霉菌、小核菌、轮枝菌等经济危害性突出的土传致病菌,显著减轻各类果蔬作物对应的真菌病害。该技术对真菌病害的防控持效期普遍可达两个生长季,部分场景下有效期更长。
土壤太阳能消毒可有效防治农杆菌、密执安棒形杆菌、梨火疫病菌诱发的细菌性病害,但难以降低青枯雷尔氏菌造成的番茄青枯病发病程度。相较于真菌与杂草,该技术对植物寄生线虫的防效偏弱,原因在于线虫较病原菌、杂草更快重新侵染定植土壤;但大田与温室试验结果证实,太阳能消毒能够稳定减轻根结线虫(根结线虫属)的根系危害、压低虫口基数,对马铃薯金线虫、胡萝卜胞囊线虫等胞囊线虫以及洋葱茎线虫也能实现理想防控。
杂草受太阳能加温的防控效果因杂草类型而异:一年生杂草基本可被彻底灭杀,多年生杂草因深层繁殖器官避开致死高温环境,防控难度更大;太阳能消毒对杂草的后效作用远优于对线虫与多数病原真菌的持效效果。
在有机农业生产、城郊农田种植、登记可用农药稀缺的特色作物种植等限药、禁药种植场景中,土壤太阳能消毒技术适用性极强。多项投入产出对比分析表明,该技术经济效益可观,还具备操作简便、适配多种耕作模式、可与其他植保措施配套联用的优势,完全契合可持续农业所倡导的有害生物综合治理理念。
9.1 引言
过去数十年间,土壤施用型农药已被广泛用于绝大多数蔬果作物,有效防治土传病害、田间杂草与植物线虫。但这类农药对人畜存在毒性风险,同时伴随高昂的环境治理成本与经济投入(皮门特尔等人,1992;鲁佐,2006),引发各界对环境安全与人体健康的深度担忧。这一现状促使应用范围最广、防效最优的化学药剂 —— 溴甲烷逐步被淘汰(卢肯、格罗夫,2006),各类在用农药的施用规范也日趋严苛(珀金斯、帕特森,1997)。
化学农药可用品类不断受限,倒逼行业转向低农药投入或无化学药剂的病虫害防控技术,也让农户与科研人员重新重视土壤太阳能消毒。该技术操作简便、环境友好,是实用性突出的非化学防控手段。
早在 20 世纪 60 年代初,农田地膜覆盖就作为农艺措施投入应用,能够减少土壤水分蒸发、防控水土流失并改良土壤理化性状(瓦戈纳等人,1960;伯罗斯、拉尔森,1962;赖,1974);而利用太阳能产生的高温灭杀土壤与植株病源生物的相关研究,面世时间更早(格罗舍沃伊,1939;纽霍尔,1955)。不过现代定义上的土壤太阳能消毒,即借助太阳能实现土壤灭除土传病原菌与杂草的专用技术,诞生于 20 世纪 70 年代以色列与约旦开展的系列研究(卡坦等人,1976)。
卡坦等人的研究成果发表后的首个十年里,全球大量科研围绕太阳能消毒技术展开,验证其对各类蔬菜、大田作物、观赏植物及果树的土传病原物、杂草与土壤节肢动物的防控效果。科研人员还系统探究了该技术的物理、化学、生物学作用原理,以及消毒处理期间和处理后土壤随之发生的各类伴生理化、生物变化(卡坦等人,1987;德韦,1991)(图 9.1)。
相关研究最初集中在地中海沿岸与中东国家,包括以色列(格林斯坦、奥舍,1991)、意大利(加里巴尔迪、古利诺,1991)、西班牙(贝略等人,2001)、希腊(亚莫斯等人,2000)与美国加利福尼亚州(斯泰普尔顿、德韦,1986);后续温带冷凉气候区等多国也陆续发布相关试验成果(加里巴尔迪、塔米耶蒂,1984)。试验场景兼顾露地与设施温室栽培体系,技术应用还拓展至育苗床、盆栽基质、冷棚苗床的土壤消毒工作(斯泰普尔顿,2000)。
随着农作物有害生物综合治理理念不断落地普及(科根,1998),此后数十年的相关研究不再将土壤太阳能消毒视作单一独立防控方案,而是将其纳入复合型病虫害防治体系的组成环节(斯泰普尔顿、德韦,1995;谢莱米,1998)。研究方向聚焦于搭配其他防控手段,强化太阳能加温的消杀效果,补齐该技术在应用与经济性上的短板(西科拉等人,2005)。

图 9.1 意大利南部田间试验中,聚乙烯地膜覆盖开展土壤太阳能消毒的试验田(资料由尼古拉格雷科博士提供)
简单来讲,土壤太阳能消毒是在环境高温时段,用地膜密闭土壤表层、截留太阳辐射热能,提升表层土温至可致死或亚致死土传病原菌与杂草的处理工艺。尽管工艺看似简易,现有研究证实其防控效果由多重复杂机理共同驱动,同时受土壤温度、地膜材质等大量环境与工艺参数制约。除灭杀土传病虫杂草外,已有大量文献证实太阳能消毒会诱发土壤理化与微生物群落的系列改变,进而改良土壤性状、影响作物产量。
本章依托截至 2008 年年初的现有文献,系统综述土壤太阳能消毒的各项研究进展:从作用机理、影响因子,到药剂靶标生物防控效果与土壤性状改良作用;同时结合可持续农业的核心支撑 —— 有害生物综合治理,汇总太阳能消毒与其他防控技术联用的现有研究成果。
9.2 土壤太阳能消毒作用机理
现有文献记载了土壤太阳能消毒繁复多样的作用机制,由多项相互关联的生理生化、生态过程协同构成(卡坦,1987;斯泰普尔顿、德韦,1995;斯泰普尔顿,1997),但其核心防控作用主要来源于太阳能带来的高温胁迫,以及高温诱导产生的土壤化学、微生物群落变化。
9.2.1 热作用机理
土壤太阳能消毒处理能够提升土体温度,达到可杀灭多数植物病原物与有害害虫的致死温度区间,因此直接高温灭活是该技术最核心、也是最常规的作用机理。部分围绕生物高温敏感生化机理的研究提出:生物的耐热差异性和细胞内大分子结构细微差异相关,高温会造成分子内氢键、离子键、二硫键异常增多并最终引发细胞死亡(布罗克,1978)。桑德拉姆(1986)则提出,温度突破细胞膜临界流动性上限后,细胞膜生理功能受损;中温型微生物(含土传病原菌与害虫)细胞膜富含低熔点不饱和脂肪酸,因此对高温耐受性极差,而耐热、嗜热型土栖生物依靠大分子在 60℃条件下仍能维持结构稳定,可在太阳能消毒处理中存活。另有研究证实,高温致使生物呼吸酶失活,是土栖微生物与杂草种子受热衰亡的另一重要诱因(布罗克,1978;桑德拉姆,1986)。
普尔曼等人(1981)提出,太阳能消毒对微生物、杂草的高温灭杀效应遵循土壤温度与受热时长呈对数负相关规律,该相关系数随靶标生物种类不同存在明显区别。绝大多数中温型土壤生物的受热损伤临界温度在 39~40℃,而田间太阳能消毒的常规升温条件无法致死嗜热、耐热型生物(斯台普莱顿、德维,1995)。科研人员已通过试验测定了多种重要致病真菌、线虫及杂草的高温灭活临界参数。
实验室条件下,波特与梅里曼(1983)测得尖孢镰刀菌、不规则腐霉、芸苔根肿菌、白腐小核菌、齐整小核菌、小核盘菌、大丽轮枝菌等多种病原真菌的致死温度区间为 38~55℃;其中尖孢镰刀菌、不规则腐霉、芸苔根肿菌耐热性最强,大丽轮枝菌、白腐小核菌、小核盘菌对高温最为敏感。大丽轮枝菌 90% 个体致死剂量(ED₉₀)条件为:37℃持续处理 14 小时,升温至 50℃时仅需 9 分钟(普尔曼等,1981a);褐座坚壳菌(异名:褐纹丝核菌)耐热能力极弱,38℃恒温处理 4 小时即可实现 50%~100% 菌体死亡(斯泰因伯格等,1987)。肉桂疫霉、草莓疫霉、巨孢疫霉在 45℃土壤中分别处理 20 分钟、30 分钟、30 分钟即可全部灭活(华雷斯 - 帕拉西奥斯等,1991);也有文献记载,38~40℃环境持续 1~2 小时可致死肉桂疫霉菌丝体(加洛等,2007)。另有试验显示:肉桂疫霉厚垣孢子悬浮于 38℃清水仅 30 分钟便全部失活(巴伯切克、冯・布龙布森,1986);带菌小麦籽粒浸泡在 41℃水中 25 分钟后病原菌被杀灭(塞隆等,1982),这也说明水的热传导效率显著高于土壤与琼脂培养基(本森,1978;科埃略等,2001)。菜豆炭腐病菌、瓜果腐霉等其余土传致病真菌耐热性更强:高含水量土壤中,40℃处理 24 小时可大幅削减菜豆炭腐病菌微菌核数量,但需 50℃及以上高温才能彻底灭活(米海尔、奥尔康,1984;谢赫、加法尔,1987)。
普尔曼等(1981a)研究表明:随处理温度升高、受热时间延长,有害生物受热损伤逐步累积直至彻底失活,亚致死高温同样会对靶标生物产生不可逆伤害。已有研究证实,真菌在亚致死温度胁迫下会合成热休克蛋白以提升自身耐热能力(普莱索夫斯基 - 维格、布兰布尔,1985;林德奎斯特,1986;弗里曼等,1989)。亚致死高温虽不能直接杀死病原菌,但会损伤繁殖体,降低孢子萌发能力与侵染致病性,同时提升病原菌受其他生物、非生物胁迫致死的概率(弗里曼、卡坦,1988;德维、卡坦,1991)。经亚致死高温弱化后的齐整小核菌菌核极易被罗氏木霉等有益微生物定殖侵染(利夫希茨等,1983;格林伯格等,1984);实验室试验中,亚致死预处理后的褐座坚壳菌繁殖体被木霉侵染的风险大幅上升(斯泰因伯格等,1987)。田间试验进一步证实:短周期太阳能消毒带来的亚致死高温,搭配减量溴甲烷或威百亩药剂,可高效防控齐整小核菌、罗勒专化型尖孢镰刀菌、甜瓜专化型尖孢镰刀菌、番茄生根专化型尖孢镰刀菌(埃谢尔等,2000)。亚致死高温对病原菌的抑制作用,也被认为是经太阳能消毒后的土壤抑病能力普遍更强的关键原因之一(格林伯格等,1987)。
环境温度持续高于 35℃,会直接致死植物寄生线虫,或破坏线虫侵染能力、提升线虫受各类生物与非生物胁迫的敏感度(希尔德、罗宾逊,1987);但太阳能消毒的杀线效果存在极强物种特异性(格雷科等,1998;达达博等,2005)。吉布林 - 戴维斯与韦尔凯德(1988)发现,带土在 48±2℃恒温下放置 2 小时,可彻底灭杀长尾刺线虫、矛形纽带线虫。大豆胞囊线虫胞囊内幼虫在 63℃仅 1 秒、52℃持续 8 分钟、44℃持续 8 小时全部死亡(远藤,1962);60℃恒温处理 30 分钟能致死马铃薯金线虫(埃文斯,1991)。沃克(1962)确定根结线虫虫卵与卵块致死临界条件为 48℃保温 6 分钟,该结论与戈克特、马图尔(1995)试验结果一致:葡萄砧木经 48~53℃热水处理 10~20 分钟,可根除根结线虫。近代试验测得南方根结线虫 95% 致死剂量(LD₉₅):39℃需 813 分钟、42℃需 281 分钟、46℃仅需 32.4 分钟(鲁伊斯等,2003)。
不同杂草种子的高温致死所需温时组合差异悬殊。埃格利(1990)测定 8 种常见杂草种子半数致死(LD₅₀)条件为 50~66℃持续 12 小时;霍洛维茨等人(1983)也证实土壤温度稳定超 45℃即可实现良好除草效果。达尔奎斯特等(2007)的室内试验显示:受试所有杂草种子在 50℃及以上高温全部死亡;马齿苋耐受上限 46℃、白苋耐受上限 42℃、稗草与龙葵耐受上限 39℃,低于对应温度加热无明显灭活作用。香附子耐热性最优:块茎在 30~90℃区间恒温处理 30 分钟,活性随温度升高呈线性下降规律(鲁宾、本杰明,1984)。科研人员还构建模拟模型,依托热处理时长或土壤昼夜变温数据预测杂草死亡率,以此推算田间太阳能消毒高效除草所需的温度 - 时长配比(达尔奎斯特等,2007;迈尔斯等,2002)。
9.2.2 化学作用机理
土壤经日光消毒(太阳能土壤热处理)后发生的各类化学变化,是日光消毒产生防控效果的另一类作用机制(陈、卡坦,1980;陈等,1991)。多数研究发现,经过日光消毒处理的土壤可溶性矿质养分含量普遍上升(卡坦,1987;斯泰普尔顿、德韦,1995;斯泰普尔顿,2000),核心诱因是热处理杀灭土壤微生物,微生物死亡后发生降解分解释放养分。在高有机质、高含水量土壤中,硝化微生物受热死亡造成土壤微厌氧环境,进而导致有毒氨态氮在土体中累积(哈松等,1987);而有机质匮乏的土壤,若日光覆膜增温效果差、土壤含水量偏低,土壤生物存活率更高、土壤通气条件好,含氮化合物可快速硝化,因此这类土壤中易硝化氮素的释放量极低(斯泰普尔顿等,1985)。此外,日光消毒带来的高温会促使土壤有机质分解,生成对土壤生物有毒的挥发性物质,同样是消毒起效的化学途径(加姆利埃尔等,2000)。
9.2.3 生物作用机理
日光消毒处理后的土壤中,能够拮抗植物病原物的有益微生物数量显著提升。究其原因,中温型土著微生物大量受热死亡后,土壤底物与可利用养分大幅增多,为拮抗菌大量增殖创造了条件(斯泰普尔顿,1981;加姆利埃尔等,1989;斯泰普尔顿、德韦,1995)。保罗与克拉克(1996)提出假说:高温分解释放的养分可提升拮抗细菌的养分同化效率,助力其在土壤菌群竞争中占据优势。除此之外,古普塔与耶茨(1997)认为,日光消毒定向灭杀取食细菌的原生动物与捕食型生物,大幅降低了有益微生物的被捕食压力,进一步推动土壤微生物群落结构发生有益转变。
芽孢杆菌、荧光假单胞菌、耐热真菌以及部分自由生活型线虫等有益拮抗生物,可耐受日光消毒高温,或是在处理结束后快速定殖土壤,既能阻止有害病虫再度侵染定植,还可促进作物健壮生长(卡坦,1987;加姆利埃尔、斯泰普尔顿,1993a;斯泰普尔顿、德韦,1995)。另有研究证实,日光消毒后土壤腐殖质含量上升,可正向促进荧光假单胞菌繁殖(陈等,1994、2000)。在经受日光高温处理后存活的微生物里,木霉属耐热真菌能够抑制多种病原真菌生长、减轻土传真菌病害发生(本 - 耶佩特等,1987;哈曼,2000;苏亚雷斯等,2004)。
9.3 影响日光土壤消毒效果的关键因素
日光消毒效果由多项因子共同决定,学界普遍认为:土壤温度、土壤含水量、气候气象条件、覆盖地膜的材质与理化特性是决定日光消毒成效的核心要素(卡坦等,1987;斯泰普尔顿、德韦,1995)。
9.3.1 土壤温度
土壤温度是决定日光热处理效果的首要指标,绝大多数中温生物的临界致死温度约为 37℃,土壤积温突破该阈值后才能发挥灭杀作用(前述文献)。斯泰普尔顿(1997)观测发现:日光消毒期间,白昼土层近地表温度最高,随土层加深、入夜降温,土体温度逐步下降。铺设透明地膜后,仅表层 0~5 cm 土壤温度可突破 50℃;温暖地区夏季开展日光消毒时,10~15 cm 土层温度可达 40~50℃,20~30 cm 土层维持在 36~40℃,更深土层升温幅度仅 3~4℃,通常达不到有害生物的亚致死或致死温度(波特、梅里曼,1983;斯泰普尔顿、德韦,1983;格雷科等,1985;切莱米等,1994)。斯泰普尔顿与德韦(1986)研究发现 46~91 cm 深层土壤线虫种群数量也出现下降,推测深层线虫消亡源于高温产生的挥发性有毒物质等其他抑害因子。相较露地大田,密闭温室环境或盆栽基质经过覆膜日光处理后,土体升温幅度更高(卡尔蒂亚,1998;斯泰普尔顿等,2000;卡斯特鲁奥沃等,2005)。
现有研究已开发多款数值模型,用于预测裸地与覆膜土壤在日光消毒过程中的温度变化(马赫尔,1979、1980)。马赫尔与卡坦(1981)、马赫尔等(1984)构建一、二维数值模型,依托环境气象数据、土壤物理性状、地膜光学参数模拟消毒土壤的温湿度变化,但该模型需要稀缺的实测气象资料,落地应用受限。基于同类理论,滕贝赫(1990)、霍顿与钟(1991)采用太阳辐射、气温、风速、降雨量等易获取气象数据,建立裸土温度预测模型。布里斯托与坎贝尔(1986)的一维模型可同步模拟土壤热量与水分迁移规律,但未纳入地膜厚度、铺设方式带来的影响。塞尼斯(1989)构建点位专属模型,模拟均质土壤日内正弦式温度波动规律;隋等(1992)的模型可测算不同覆膜条件下土壤温湿度垂直分布。还有部分模型聚焦地膜光学性能对土壤增温的调控作用(哈姆、克洛伊滕贝格,1994;吴等,1996;鲁奥科,2000);格雷费(2005)建立能量平衡模型,可适配垄面局部覆膜的二维地表温度测算。
9.3.2 土壤含水量
土壤含水量同样是调控日光消毒热效应的关键因子:水分可大幅提升热量向土壤微生物、杂草种子与作物残体的传导效率;同时,适宜水分能激活种子生理活性、促进土栖微生物代谢增殖,使其对高温胁迫更加敏感,更易被高温灭杀。
模型模拟结果表明,土壤含水量提升有助于土层达到峰值温度(马赫尔等,1984;诺特等,1987),也有研究证实土壤热容量与含水率呈线性正相关(德弗里斯,1963;塞斯韦伦等,2006)。但阿尔 - 卡拉古利与阿尔 - 凯西(2001)得出相反结论:土壤最高温度随含水量升高而降低,据此提出日光消毒期间无需反复灌水即可实现土传病原消杀。
9.3.3 气候与气象条件
气候和天气直接决定地表太阳辐射强度,进而左右土壤升温幅度,是日光消毒成败的先决条件。切莱米等(1997)试验表明,阴雨多云天气会大幅削弱日光消毒效率;总体来看,夏季高温区域采用该项技术防控效果最优。不过,温暖区域夏季虽是日光消毒的最佳窗口期,但已有大量文献证实,在冷凉气候区,日光覆膜处理同样可有效防治土传病原、线虫、细菌性病害与田间杂草(拉约等,1997;克里斯滕森、辛戈德,1999;平克顿等,2000;皮奇等,2001;塔米耶蒂、瓦伦蒂诺,2006)。冷凉区域也能实现良好防控的原因在于:病原菌耐热阈值随环境气候发生适应性改变,冷凉环境下病原致死临界温度更低(埃尔莫尔,1998)。
注释说明 1.soil solarization:农学通用译名为日光土壤消毒 / 太阳能覆膜土壤热处理(行业简称日光消毒),是田间覆透明地膜依靠太阳能升温灭菌的农法; 2.外文人名、年份保留原文标注格式,符合农学论文参考文献规范; 3.ammoniacal nitrogen:氨态氮;fluorescent pseudomonads:荧光假单胞菌;Trichoderma spp.:木霉属,均采用国内土壤植保标准译名。 |
9.3.4 塑料薄膜
日光土壤消毒工艺中,塑料薄膜的作用是透过太阳辐射提升地温,同时减少辐射散热与对流散热损失(帕帕达基斯等人,2000)。塑料薄膜对太阳辐射的透过能力与其辐射光学性能密切相关,核心指标为透光率:薄膜太阳透光系数越高,膜下地温增幅越显著(斯卡拉西亚 - 穆尼奥扎等人,2004;沃克斯等人,2005)。除此之外,反射率、吸收率、发射率等其他辐射光学参数同样会影响塑料薄膜的升温效果(帕帕达基斯等人,2000)。科研人员已通过室内试验与田间试验,系统测定了各类塑料薄膜的辐射光学特性(皮尔逊等人,1995;帕帕达基斯等人,2000;海斯纳等人,2005;沃克斯等人,2005)(图 9.2)。
已有大量研究证实多种塑料原料均可用于日光土壤消毒(兰贝蒂、巴锡莱,1991),其中聚乙烯(低密度聚乙烯、乙烯 - 醋酸乙烯酯共挤改性聚乙烯,可单独使用或添加无机盐助剂)、乙烯 - 醋酸乙烯酯共聚物与聚氯乙烯薄膜的日光升温综合表现最优(史蒂文斯等人,1991a)。针对不同薄膜辐射光学性能开展的室内与田间对比试验表明,乙烯 - 四氟乙烯共聚薄膜的分光辐射性能优于乙烯 - 乙烯醇共聚物膜、聚乙烯膜、红光选择性滤光膜及紫外阻隔膜,能够实现更高的土壤升温幅度,提升日光土壤消毒成效(卡斯科内等人,2005)。
温室栽培环境下,采用乙烯 - 四氟乙烯共聚薄膜进行日光土壤消毒,尽管土壤升温上限不及乙烯 - 醋酸乙烯酯薄膜,却能更高效防控多种重要土传病原物(波利齐等人,2003)。马拉斯拉基斯与卢拉基斯(1989)研究显示:聚乙烯薄膜与聚偏二氯乙烯薄膜对齐整小核菌的防控效果相近,但只有聚偏二氯乙烯薄膜可显著减轻由顶孢霉引发的黄瓜根腐病与维管束褐变;蔡斯等人(1999a)发现红外高吸热透明薄膜提升地温的效果持续优于低密度聚乙烯膜。阿尔 - 凯西与阿尔 - 卡拉古里(2002)通过数值模拟与田间试验证实,相较于普通透明聚乙烯地膜,石蜡乳液成膜覆盖可更高效加热土壤、快速灭杀土传致病真菌。
聚乙烯凭借优异的太阳光透过性、柔韧性、拉伸强度与耐候性,普遍被视作日光土壤消毒的优选覆膜原料(埃斯皮等人,2006)。薄型聚乙烯地膜性价比更高,在地温提升效果上优于厚膜(斯台普莱顿、德维,1986;阿布 - 伊尔迈莱,1991a、b)。双层聚乙烯覆膜比单层覆膜升温效果更好,可使地温提升 3~10℃,强化对靶标有害生物与土壤微生物群落的调控作用(本 - 耶佩特等人,1987;塞尼斯,1987;马哈茂德,1996),尤其适用于育苗圃以及多云寡照区域(安内西、莫塔,1994;史蒂文斯等人,1999;罗德里格斯佩雷斯等人,2004)。

图 9.2 意大利南部塑料大棚内不同薄膜日光土壤消毒试验(图片由多纳托卡斯特鲁奥沃博士提供)
众多学者还围绕地膜颜色开展相关研究,地膜颜色直接决定覆膜的能量辐射特性,进而影响土壤含水量与作物根区微环境(拉蒙特,1993;斯特雷克等人,1995;埃尔 - 凯布劳伊等人,2006)。海恩斯(1987)指出,不同颜色地膜对地温的提升能力排序通常为:透明膜>黑膜>白膜。巴拉卡特(1987)试验表明,黑色不透明膜会阻隔绝大部分太阳辐射,覆膜地温较透明膜低数摄氏度;但也有试验得出透明膜与黑膜下地温变化规律相近的结论(哈辛等人,2004)。透明膜覆盖下土壤高温持续时间更长,而黑色聚乙烯地膜田间耐用性更好、控草能力更强(杜波依斯,1978;汉考克,1988;哈辛等人,2004)。国内外大量试验分别采用透明、黑色及其他配色地膜开展日光消毒,试验结果参差不齐(卡德曼 - 扎哈维等人,1986;阿布 - 加尔比赫等人,1991;哈姆等人,1993;蔡斯等人,1999;坎皮利亚等人,2000;里格等人,2001;哈辛等人,2004)。育苗床与成龄果园等特殊种植场景推荐选用黑色地膜(斯台普莱顿、加尔萨 - 洛佩斯,1988;阿布 - 加尔比赫等人,1991;斯台普莱顿,1997)。新建果园树盘覆盖黑色聚乙烯地膜,相较无覆膜空白对照,杂草发生量与根结线虫根瘤发生率均明显下降(邓肯等人,1992)。部分研究表明黑膜日光消毒能够压低多种土传致病真菌、细菌种群基数,除草效果突出(雷诺兹,1970;霍索恩,1975;拉蒙特,1993);也有试验显示:黑膜可促进作物早熟增产,但全年总产量低于透明地膜栽培(肖恩贝克、埃瓦尼奥,1998)。
使用塑料薄膜实施日光土壤消毒,需要配套专用机具、耗费大量人工工时,作物采收后废旧地膜的回收处置成本高昂,还会带来突出的环境污染问题(帕里什等人,2000)。现有多项研究依托高分子材料改良技术,探索上述难题的解决方案(施派希等人,1990;多兰,2002;郑等人,2005)。按适宜用量喷施于土表的可喷涂降解高分子成膜材料,是替代传统塑料地膜开展日光土壤消毒的经济可行方案(加姆利埃尔等人,2001)。尽管喷施降解膜的地温峰值普遍低于常规塑料地膜,但仍可有效防治马铃薯疮痂病、花生果腐病,同时提升茄子单果重与总产量(马哈茂德普尔、斯台普莱顿,1997;加姆利埃尔等人,2001)。
以可再生农业原料制备的生物可降解地膜,因可就地入土降解或送入堆肥厂资源化处理,成为传统塑料地膜的环保替代方案(钱德拉、鲁斯特吉,1998;纳拉扬,2001;马林科尼科等人,2002;基里库,2007)(图 9.3)。学界针对淀粉基可降解地膜的田间降解规律与物理形态变化已开展大量研究(巴斯蒂奥利,1998;布里阿苏利斯,2004;海斯纳等人,2005;沃克斯等人,2005;斯卡拉西亚 - 穆尼奥扎等人,2006)。相较于低密度聚乙烯、乙烯 - 醋酸乙烯酯传统地膜,淀粉基可降解膜所能达到的最高地温偏低、高温维持时间更短(坎迪多等人,2005;鲁索等人,2005),但大量田间实测证实该类地膜在多种作物日光消毒中应用成效优良(钱德拉、鲁斯特吉,1998;马内拉等人,2002)。温室与露地栽培条件下,玉米淀粉基生物降解膜日光土壤消毒,可有效防控甜瓜根结线虫、番茄斑裂壳孢引发的木栓根病、生菜小核菌病害及田间杂草,同步实现提质增产(帕纳托尼等人,2004;卡斯科内等人,2005;卡斯特鲁奥沃等人,2005)。

图 9.3 意大利南部田间试验中,日光土壤消毒结束后生物降解地膜的自然降解状态(图片由多纳托卡斯特鲁奥沃博士提供)
9.4 土壤太阳能消毒的作用效果
9.4.1 对土壤理化性状的影响
无论是大田实地处理,还是人工培养箱模拟土壤太阳能消毒试验,多数研究均证实日光加温可提升土壤可溶性养分含量,尤其能提高溶解性有机质、无机态氮以及速效阳离子的含量(斯台普莱顿等,1985;史蒂文斯等,1991a;格伦茨维希等,1999;陈等,2000;萨莱诺等,2000;吉尼等,2003)。陈与卡坦(1980)检测发现,经太阳能消毒后的土壤饱和浸提液中溶解性有机质浓度上升;盖尔索米诺等人(2006)后续提出假说:太阳能消毒产生的高温促使土壤有机质发生温和水解与解聚反应,是可溶性有机质含量升高的诱因。
太阳能消毒处理后,土壤速效态氮(铵态氮\(\boldsymbol{NH_4^+}\)、硝态氮\(\boldsymbol{NO_3^-}\))短期有效性普遍提升,根源在于有机质分解速率加快,同时高温杀灭的土壤微生物残体发生矿化作用(陈、卡坦,1980;斯台普莱顿等,1985;考鲁昂等,1989a、b;艾哈迈德等,1996;格伦茨维希等,1998;弗雷塔斯等,2000;毛罗米卡莱等,2005a、b)。土壤不同形态氮素的配比变化和土壤 pH、硝化微生物数量密切相关:太阳能消毒的高温会灭杀硝化细菌,促使土壤铵态氮富集;若局部土温偏低、土壤有机质匮乏,硝化微生物可存活,生成的硝态氮极易随水淋溶流失,最终造成土壤氮素损耗(哈森等,1977;考鲁昂等,1989a)。
绝大多数研究表明,太阳能加温处理很难提高土壤全磷含量(陈、卡坦,1980;斯台普莱顿等,1985;考鲁昂等,1989b;陈等,1991),仅有少量文献报道该处理可提升土壤总磷或水溶性磷的有效性(考鲁昂等,1989a;盖尔索米诺等,2006)。钾、钙、镁、钠等阳离子经太阳能消毒后,土壤速效含量整体呈上升趋势(陈、卡坦,1980;斯台普莱顿等,1985;考鲁昂等,1989b;加姆利埃尔、卡坦,1991;艾哈迈德等,1996;格伦茨维希等,1998)。
几乎所有相关试验都观测到太阳能消毒能促进作物长势,核心原因就是上述大量中微量元素有效性提升,同时腐殖质溶解活化微量养分,利于作物吸收利用(陈、阿维亚德,1990;陈等,1991)。鉴于太阳能消毒对土壤养分的活化增效作用,弗洛雷斯等人(2007)提出建议:土壤日光加温前应减少化肥施用,避免作物营养过盛造成茎叶徒长、产量下降。
多项研究测得太阳能消毒后土壤溶液电导率上升(陈、卡坦,1980;斯台普莱顿等,1985;考鲁昂等,1989a;艾哈迈德等,1996),现有推论认为:有机质分解矿化释放大量离子,从下层土体向上迁移汇集至受热表层土壤溶液,进而抬升电导率(陈、卡坦,1980)。另有观点指出,昼夜温差驱动土壤水分与可溶性盐分向下淋洗,这也是部分试验中太阳能消毒可降低土壤含盐量的关键机理(阿卜杜勒 - 拉希姆等,1988;阿尔 - 凯西等,1989)。高温处理对土壤导水率的影响结论不一,不同试验结果存在显著分歧(陈、卡坦,1980;阿尔 - 凯西等,1989)。总体来看,太阳能消毒对土壤物理性状影响幅度有限且效果不稳定(陈等,1991),但梅莱罗 - 瓦拉等人(1989)的试验证实,该处理能够改良土壤结构、促进土壤团粒形成。
9.4.2 对非致病性土壤微生物区系的影响
土壤太阳能消毒除灭杀多数植物病原微生物外,还会广泛扰动土壤各类有益微生物群落;现有研究共识为:加温处理显著改变土壤微生物群落组成与物种丰富度(陈等,1991;舍恩菲尔德等,2003;帕莱塞等,2004;卡尔曼等,2006;盖尔索米诺等,2007)。阿拉布韦特等人(1996)提出假说:高温打破原有土壤微生物平衡,腐生型微生物获得生长优势,加剧土壤养分种间竞争,最终提升土壤抑病能力。近期温室太阳能消毒试验表明,加温会降低土壤微生物生物量与土壤酶活性,而土壤有机质可缓冲高温对微生物的伤害,起到保护作用(奥库尔等,2006;斯科帕、迪蒙泰,2007)。
部分文献记录太阳能消毒造成土壤细菌总数量下降(马哈茂德,1996;帕特尔夫妇,1997;伊藤等,2000;巴伯等,2002;夏尔马等,2002);也有研究发现处理后土壤真菌数量减少,但细菌种群不受明显影响(科茨 - 贝克福德等,1997;舒克拉等,2000)。现有针对土壤细菌的相关研究,大多聚焦促生菌、病原菌拮抗菌,以及可在消毒土壤根系快速定殖的土著菌群(斯台普莱顿、德维,1984;史蒂文斯等,1991b;瓦迪,1999)。其中部分试验结果显示,太阳能消毒土壤中细菌与放线菌总量不降反升(考鲁昂等,1989;海尔、巴基尔,1995;卡莱克等,1999)。史蒂文斯等人(2003)发现,消毒后土壤细菌群落结构发生演替,根际细菌、芽孢杆菌、荧光假单胞菌成为优势类群;而斯台普莱顿与德维(1982、1984)的试验得到相反结果:荧光假单胞菌、芽孢杆菌、放线菌、农杆菌经高温处理后种群密度大幅下降。里斯泰诺等(1991)、加姆利埃尔与卡坦(1991)同样观测到多数土壤细菌被抑制,放线菌受高温影响相对更小。
荧光假单胞菌具备极强的土壤快速复殖能力(斯台普莱顿、德维,1982、1986;加姆利埃尔等,1987),既能防控土传真菌性根部病害,又可促进作物生长(利夫希茨等,1983;格林伯格等,1984;斯台普莱顿、德维,1984;弗里曼、卡坦,1988;托马肖、韦勒,1990;基尔,1992;陈等,2000)。大量研究证实,芽孢杆菌是太阳能消毒后土壤中主要存活的革兰氏阳性菌,凭借生长速度快、可分泌抑菌抗生素两大特性,成为消毒土壤抑病功能的关键菌群(斯台普莱顿、德维,1982、1984;卡坦,1987)。
科研人员从经太阳能消毒的番茄根尖筛选得到根内生芽孢杆菌菌株,离体试验证实该菌株可高效抑制大丽轮枝菌菌丝生长,田间试验能有效防控茄科作物黄萎病(提亚莫斯、帕普洛马塔斯,1987;提亚莫斯等,2004)。此外,大丽轮枝菌另一类重要拮抗菌 —— 黄孢原毛平革菌,同样耐受太阳能高温处理,且可在作物根际大量增殖(金等,1988;提亚莫斯、弗拉韦尔,1995)。
因根瘤菌在生物固氮体系中的关键作用,诸多试验专门探究太阳能消毒对土壤根瘤菌群的影响。多数结果表明:高温降低土壤土著根瘤菌丰度,抑制苗期作物根系结瘤(阿卜杜勒 - 拉希姆等,1988;乔汉等,1988;林克等,1991;马哈茂德,1996),但豆科作物定植后,土壤根瘤菌种群可快速恢复(林克等,1991)。毛罗米卡莱等(2005a、b)发现,蚕豆、鹰嘴豆经消毒土壤栽培后根系结瘤推迟、单株根瘤数量显著减少;也有学者提出,利用太阳能消毒杀灭本土根瘤菌,便于人工接种优良商品根瘤菌剂实现菌种替换(鲁佩拉、苏达尔沙纳,1990)。不过阿罗拉与潘迪(1989)、奈尔等(1990)的试验得出不同结论:太阳能消毒反而提升了土壤根瘤的结瘤效率。
目前关于太阳能土壤消毒对丛枝菌根真菌(AMF)产生副作用的相关研究数量稀少且结论相互矛盾,但多数研究结果表明土壤升温处理不会损伤本土丛枝菌根真菌,反而能够提升植物根系菌根侵染率与植株长势(普尔曼等人,1981;阿费克等人,1991)。达夫特等人(1987)研究发现,明亮球囊霉孢子在 45℃条件下失活;门格等人(1979)则报道,聚丛球囊霉在 51.5℃环境下处理 10 分钟即可受热死亡。苏拉什等人(1997)观测到,外生菌根真菌在环境温度超过 45℃时生长受到抑制,是土传真菌中对太阳能升温最敏感的类群,并据此提出:太阳能土壤消毒可作为林业育苗圃可控人工菌根培育的有效土壤消杀手段。斯塔普莱顿与德韦(1984)对比太阳能消毒地块与未处理地块的作物根系,发现两类土壤中丛枝菌根真菌的根系内生侵染程度无明显差异;但另有试验显示,经过 8 周太阳能覆膜处理后,土壤土著丛枝菌根真菌种群数量降至检出限以下,而人工接种的根内球囊霉仍可保持活性(本达维德 - 瓦尔等人,1997)。坎普鲁维等人(2007)证实,太阳能土壤消毒仅能削减丛枝菌根真菌繁殖体数量,无法实现彻底根除;其中根内球囊霉接种物在 50℃高温处理后,完全丧失形成菌根的侵染能力。施赖纳等人(2001)提出假说:太阳能消毒后土壤丛枝菌根真菌丰度下降,间接诱因是田间杂草被灭杀,而杂草原本可在越冬期维系土壤菌根菌群存续。
9.4.3 对病原真菌的作用
土壤太阳能消毒对土传病原真菌的抑制效果,通常受气候条件、土壤理化性状、土壤内真菌繁殖体的种群数量与空间分布等多重因素共同影响(普尔曼等人,1979)。不过已有大量研究证实,日光加温处理对多数具备重大经济危害性的植物病原真菌均可起到理想的杀菌效果。
国内外已围绕多种作物、在不同试验环境下系统开展土壤太阳能消毒防控镰孢枯萎病的相关研究。加姆利埃尔与卡坦(1993)提出假说:除高温直接作用于病原菌造成死亡外,荧光假单胞菌等有益微生物群落也参与介导了土壤消毒带来的抑病作用。尖孢镰刀菌不同专化型的耐热能力存在明显分化(什莱温等人,2004),但大田太阳能消毒仍可显著降低孜然、玉米、陆地棉、西瓜、结球甘蓝上由尖孢镰刀菌诱发的枯萎病发病概率(洛达,1995;伊斯雷尔等人,2005;艾哈迈德等人,1996;卡坦等人,1983;马丁 & 哈茨,1986;拉米雷斯 - 维拉普杜阿 & 芒内克,1987、1988)。美国佛罗里达、以色列、土耳其的田间试验均表明,土壤太阳能消毒能够有效防治番茄镰孢枯萎病(谢莱米等人,1997;加姆利埃尔 & 卡坦,1993;于塞尔 & 奇纳尔,1989),但另有研究显示,该技术仅能消减表层 5 厘米土层内的病原菌菌量(谢莱米等人,1994)。澳大利亚田间试验结果显示,太阳能消毒可降低香石竹尖孢镰刀菌的土壤接种体量,却无法减轻香石竹与西瓜田间病害表观症状(波特 & 梅里曼,1985)。马铃薯、西瓜田间试验证实,经过 30~60 天土壤太阳能消毒处理后,0~30 厘米土层内的茄腐镰刀菌种群大幅下降甚至被彻底铲除(曼苏里 & 贾利亚尼,1996;特里基等人,2001)。温室栽培条件下,太阳能消毒可高效防控香石竹尖孢镰刀菌石竹专化型病害(埃莱娜 & 贾莫斯,1992)、控制番茄镰孢枯萎病(约安努,2000);以色列试验中,该处理使番茄根腐尖孢镰刀菌种群数量下降 69%~95%(什莱温等人,2003)。此外,温室日光土壤消毒近乎完全根除西瓜尖孢镰刀菌西瓜专化型、甜瓜尖孢镰刀菌甜瓜专化型(冈萨雷斯 - 托雷斯等人,1993;塔米耶蒂 & 瓦伦蒂诺,2006)。
多项土壤太阳能消毒试验证实,该技术能有效压低疫霉属真菌土壤基数、减轻对应作物疫霉病害发生(平卡斯等人,1984;洛佩斯 - 埃雷拉等人,1997;麦戈文等人,2000;本利奥格鲁等人,2005)。平克顿等人(2002)发现,日光消毒后的土壤中樟疫霉被彻底灭活,这一结论与南非巴伯切克 & 冯・布龙布森(1986)的研究结果一致;该团队此前(2000)还报道,土壤日光处理结束 2 周、4 周后,30 厘米、45 厘米土层中已检测不到樟疫霉活性。美国加州与西班牙的大田试验表明,土壤加温处理可大幅削减甚至完全消灭草莓疫霉土壤菌源(哈茨等人,1993;波拉斯等人,2007b),经太阳能消毒的地块栽种草莓后,草莓疫霉、柑橘疫霉对草莓根系的侵染程度也显著下降(哈茨等人,1993;平克顿等人,2002)。西瓜田间消毒试验显示,0~25 厘米土层内的德雷克斯勒疫霉繁殖体经日光处理后数量锐减或完全消亡(曼苏里 & 贾利亚尼,1996);烟草疫霉、马铃薯疫霉、辣椒疫霉在 10~15 厘米土层的种群密度经处理后明显降低(谢莱米等人,1994;科埃略等人,1999、2000)。澳大利亚相关试验证实,土壤太阳能消毒防控由隐地疫霉引发的酸樱桃颈腐病,防效可维持 12 个月以上(威克斯,1988)。
土壤太阳能消毒同样被应用于油橄榄园大丽轮枝菌黄萎病防控,相关田间应用效果得到验证(贾莫斯等人,1991;洛佩斯・埃斯库德罗 & 布兰科・洛佩斯,2001)。阿什沃思 & 高纳(1982)在阿月浑子果园试验中发现,日光消毒可将 120 厘米深层土壤中的大丽轮枝菌彻底杀灭;杏树、扁桃采用该处理后,黄萎病发病株率下降 86%~100%(斯台普莱顿等人,1993)。露地蔬菜试验表明,朝鲜蓟地块经土壤太阳能消毒后,连续三茬栽培均可有效控制黄萎病(贾莫斯 & 帕普洛马斯,1988),马铃薯、茄子、棉花黄萎病也能实现稳定控害(戴维斯 & 索伦森,1986;拉扎罗维茨等人,1991;平克顿等人,2000;约安努,2001;普尔曼等人,1981b;梅莱罗 - 瓦拉等人,1995)。摩根等人(1991)证实,作物定植后地表铺设太阳能消毒地膜,仍可对成株期番茄黄萎病起到良好防效。希腊克里特岛、塞浦路斯温室番茄、意大利北部温室茄子的日光土壤消毒试验,同样取得理想的黄萎病防控效果(布尔博斯 & 斯库德里达基斯,1996;约安努,2000;塔米耶蒂 & 瓦伦蒂诺,2001)。
不同研究中,土壤太阳能消毒对齐整小核菌所致病害的防控效果虽存在差异,但整体防效均达显著水平(雷诺兹,1970;格林斯坦等人,1979a;米海尔 & 奥尔康,1984;史蒂文斯等人,1990b;里斯泰诺等人,1991;谢莱米等人,1997;什莱温等人,2003)。果园土壤日光消毒后,2.5 厘米土层内齐整小核菌菌核活性快速下降 95% 以上,土层越深处理效果越弱(斯台普莱顿等人,1989)。西班牙、新西兰的试验结果显示,土壤太阳能消毒后,诱发大蒜白腐病的葱腐核盘菌接种体量降至微量乃至无法检出(巴萨洛特 - 乌雷巴 & 梅莱罗 - 瓦拉,1993;梅莱罗 - 瓦拉等人,2000;麦克莱恩等人,2001)。大量文献证实,太阳能消毒可高效防治由小核盘菌引发的莴苣菌核猝倒病(霍索恩,1975;万纳奇等人,1988;西尼加利亚等人,2001;帕特里西奥等人,2006、2007)。以色列田间经日光土壤消毒后,核盘菌菌核死亡率超 90%(本 - 耶佩特,1988);新西兰试验中,受消毒时长与试验地块环境影响,菌核死亡率在 53%~100% 区间浮动(斯瓦米纳坦等人,1999)。菲利普斯(1990)提出观点:土壤日光加温产生的亚致死高温削弱菌核生理活性,再经由土壤微生物侵染分解,是菌核活性大幅衰退的核心诱因。
由于菜豆拟茎点霉耐热性较强,采用土壤太阳能消毒防控该病原菌需要匹配适宜的温度与湿度条件(博伦,1985;米海尔 & 奥尔康,1984;谢赫 & 加法尔,1987)。哈茨等人(1987)的大田试验中,日光消毒未能在任意土层降低菜豆拟茎点霉菌量,但后续研究发现,该处理可显著减少瓜尔豆、玉米土壤内病原菌数量,缓解作物干腐根症状(洛达,1995;哈米德 & 阿斯兰,1996;洛达等人,1997)。
众多研究证实土壤太阳能消毒对多种作物上的立枯丝核菌具备稳定抑制作用(普尔曼等人,1981;切特等人,1982;考鲁昂等人,1989a、1989b;凯纳斯,1995;卡坦,1996)。尤其对于凤仙花颈腐与枯萎、草莓和莴苣立枯病,土壤日光消毒可有效压低病害终发率、延缓病害扩展进程(麦戈文等人,2002;平克顿等人,2002;帕特里西奥等人,2006、2007)。
加那利群岛、意大利、葡萄牙温室试验表明,土壤太阳能消毒能够高效防控番茄由番茄壳二孢引发的木栓根腐病(迪亚斯・埃尔南德斯等人,2005;加里巴尔迪 & 塔米耶蒂,1984;卡尔蒂亚等人,1989;卡斯孔 & 德埃米利奥,2000;莫拉 & 帕尔米尼亚,1994);埃及沟灌农田田间试验同样收获良好防效(阿卜杜勒 - 拉希姆等人,1988),塞浦路斯四项试验仅实现部分控病(约安努,2001)。韭葱根系、洋葱幼苗及鳞茎的粉红根腐病由粉红腐病菌侵染所致,多数试验证明土壤太阳能消毒可大幅减轻该病危害(哈茨等人,1989;加姆利埃尔等人,2004;陈荣等人,2007)。
大田土壤太阳能消毒可大幅减少甚至彻底消除腐霉对胡萝卜、草莓根系的侵染(贝克尔 & 沃纳,1995;平克顿等人,2002),西瓜、马铃薯地块经处理后,25~30 厘米土层内瓜果腐霉菌量显著下降(曼苏里 & 贾利亚尼,1996;特里基等人,2001)。即便在气候温凉的丹麦,夏季温室土壤日光消毒依旧可以有效防控腐霉根腐病(克里斯滕森 & 辛加德,1999)。
研究人员还探究了土壤太阳能热处理对其他病原真菌的潜在防治效果。田间与温室试验证实,太阳能土壤消毒可高效防治十字花科根肿病致病菌 —— 芸苔根肿菌(怀特、布扎茨基,1979;堀内、堀,1983;迈尔斯等人,1983),不过该病原菌的热敏感性依赖适宜的土壤湿度环境(波特等人,1991)。在苹果与鳄梨果园(施泰因贝格等人,1987;弗里曼等人,1990;洛佩斯 - 埃雷拉等人,1998)以及苹果育苗圃(夏尔马等人,2002)中,利用太阳能土壤消毒技术可有效防控褐座坚壳菌(异名:褐丝孢束梗霉)。温室试验发现,无论土壤栽培(堀田、真部,2005)还是无土栽培体系(吉拉多等人,2005)中,甜瓜坏死斑点病毒的传毒媒介 —— 沃氏油壶菌(异名:根生油壶菌)存活率均大幅下降。在夏季高温气候的摩洛哥,温室条件下的太阳能土壤消毒技术同样能有效防治番茄茎枯病菌引发的茎溃疡病害(贝斯利,1982)。
专业名词注释(便于农学查阅) 1.soil solarization:太阳能土壤消毒(农艺通用译法,地膜覆土靠日光升温灭病虫) 2.Plasmodiophora brassicae:芸苔根肿菌,致十字花科蔬菜根肿病 3.Rosellinia necatrix:褐座坚壳菌,果树白纹羽病病原菌 4.Olpidium bornovanus:沃氏油壶菌,土壤传毒型真菌媒介 5.Didymella lycopersici:番茄茎枯病菌 |
科研人员还在育苗基质与盆栽土中探究了太阳能土壤消毒对病原真菌的抑制作用。林木苗圃相关试验表明,日光土壤加温处理在降低腐霉、镰刀菌以及立枯丝核菌的土壤侵染活性方面,效果与蒸汽灭菌、化学熏蒸相当(安内西、莫塔,1994;勒比昂等人,1997;萨莱诺等人,2000)。林木苗圃夏季太阳能土壤消毒,可显著减轻辐射松与斜叶桉种子出苗前猝倒病的发生(卡萨比,1985)。该试验同时证实,日光消毒能够降低辐射松幼苗出苗后的死苗率,还可彻底清除人工接菌松树根系中的樟疫霉、尖孢镰刀菌与各类腐霉。
考鲁昂等人(1989)研究表明,采用塑料袋密闭日光熏蒸处理受隐地疫霉、尖孢镰刀菌、立枯丝核菌污染的栽培基质,可有效防治非洲菊根腐病。杜夫与巴尔纳特(1992)研究发现,在经日光覆膜处理的堆置栽培基质中,根据病原菌种类耐热性与接种土层深度差异,瓜果腐霉、烟草疫霉以及齐整小核菌的菌源可在 3~10 天内被灭杀。对容器浅层栽培基质实施太阳能消毒,能大幅减轻由坎农单孢壳引发的甜瓜枯萎问题,还可快速降低该病菌子囊孢子的存活能力(科恩等人,2000;皮沃尼亚等人,2002)。除此之外,已有研究证实太阳能土壤消毒技术可用于废旧栽培基质的再生循环利用(齐纳蒂等人,2002)。
专业术语注解 1.containerized soil:盆栽基质 / 容器育苗土 2.preemergence damping-off:出苗前猝倒病;post-emergence mortality:出苗后幼苗枯死 3.potting mixes /growth medium:栽培基质、盆栽营养土 4.spent potting media:废弃基质、用过的栽培土 5.solarization:太阳能土壤消毒(地膜覆土日晒升温灭菌) |
大量研究表明,太阳能土壤消毒对病原真菌的防病效果通常可维持约两个生长季(乌斯玛尼、加法尔,1982;斯泰普尔顿、德韦,1986;格林伯格等人,1987;戴维斯,1991);另有试验证实,土壤经日光消毒后,土传病害的抑制效果能够持续 1~3 年(斯泰普尔顿、德韦,1982;加姆利埃尔、卡坦,1993;约安努,2000;史蒂文斯等人,2003)。该技术防治棉花镰刀菌枯萎病、花生黄萎病与果斑病、洋葱粉红根腐病以及番茄腐霉根腐病的药效,可稳定保持 2~3 个种植周期(卡坦,1981;卡坦等人,1983;拉比诺维奇等人,1985)。相关研究记录了太阳能土壤消毒对黄萎病的长效防控作用:在红花、棉花、朝鲜蓟多茬连作栽培中可长效控病(普尔曼等人,1981;亚莫斯、帕普洛马塔斯,1988),同时对番茄栓根病、洋葱白腐病菌引发的白腐病同样具备长效防治效果(阿卜杜勒 - 拉希姆等人,1988;萨图尔等人,1989)。一项为期两年的田间试验显示,土壤太阳能消毒处理后,第二茬生菜上由小核盘菌与立枯丝核菌诱发的病害发生程度大幅下降(帕特里西奥等人,2006)。
专业名词注释 1.Verticillium wilt:黄萎病 2.pink root of onion:洋葱粉红根腐病 3.corky root:番茄栓根病 4.S. cepivorum:葱腐葡萄孢(洋葱白腐病菌) 5.S. minor:小核盘菌 6.growing seasons:生长季、种植茬口 |
已有研究证实太阳能土壤消毒对林果类作物土传病原菌同样具备长效防控效果:经日光消毒处理的苹果园,连续三年未发生褐座坚壳菌诱发的白纹羽病,重栽苹果树两年内无死树现象(弗里曼等人,1990)。在油橄榄园中,热处理对大丽轮枝菌土壤菌群的抑制作用以及果树长势恢复的改善效果可维持至少三年(亚莫斯等人,1991;洛佩斯・埃斯库德罗、布兰科・洛佩斯,2001)。鳄梨果园定植前实施太阳能土壤消毒,对樟疫霉的控病时效可超过 5 年;加洛等人(2007)研究发现,即便在热处理 11 年后,处理地块植株枯死率仍显著低于未做日光消毒的对照地块。
太阳能土壤消毒对土壤病原菌的长效抑制作用无法单用高温灭菌来解释:土壤受热后,耐热性强、具备生防拮抗能力的细菌、放线菌与有益真菌会快速在土壤中定殖繁衍,进而提升土壤抑病能力,共同实现长期控病效果(亚莫斯、帕普洛马塔斯,1988;考鲁昂等人,1989;德韦,1991;加姆利埃尔、卡坦,1993;平克顿等人,2000)。
专业术语注释 1.R. necatrix:褐座坚壳菌,果树白纹羽病致病菌 2.V. dahliae:大丽轮枝菌,黄萎病病原 3.P. cinnamomi:樟疫霉,鳄梨根腐主要病原菌 4.soil suppressiveness:土壤抑病性(天然抑病土特性) 5.antagonistic microorganisms:拮抗微生物 |
4.4 对细菌与病毒的作用
有关太阳能消毒土壤中植物病原细菌存活规律的研究数量有限。农杆菌属病菌对太阳能土壤消毒高度敏感:斯泰普尔顿与德韦(1984)试验测得土壤农杆菌菌群最高降幅可达 72%;意大利两处育苗圃试验中,经日光覆膜处理后,该菌种群分别下降 99%、92%(拉约等人,1997)。该项研究结果显示,砂壤土中的两株根癌农杆菌在 4 周内被彻底灭活,粉砂黏土内的菌株则在两个月后数量大幅锐减。赫拉伊夫(2003)研究表明,土壤太阳能消毒可稳定抑制农杆菌种群、减少桃与苦杏仁砧木幼苗根癌瘤发生,但防控效果受土壤温度与土质类型制约。
番茄溃疡病病原菌 —— 密歇根棒形杆菌密歇根亚种同样具备强热敏性:什莱温等人(2004)测算得到该病菌 80% 致死所需处理时间为 7 天;安东尼乌等人(1995)发现,塑料棚内土壤覆膜日晒 4~6 周后,该病菌活菌数量急剧下降。对感染梨火疫病菌的苹果、西洋梨植株搭建棚罩实施日光加温处理,能够遏制溃疡病斑扩展,多数病树体内病菌被彻底铲除,不过防治效果随树种与环境温度变化(汤姆森,1996)。田间与塑料大棚条件下,单独采用太阳能土壤消毒无法降低青枯雷尔氏菌(异名:青枯假单胞菌)诱发的番茄细菌性病害发病率(切莱米等人,1994、1997、1999;堀内,1991),但日光消毒搭配药剂施用可有效压低土壤青枯菌基数(切莱米等人,1999)。
相较于病原菌,土壤太阳能消毒对植物病毒的影响相关研究十分匮乏,目前仅有一篇文献证实:砂壤土经热处理后,土壤中烟草花叶病毒的降解进程趋于稳定(特里奥洛、马泰拉齐,1992)。
专业名词注释 1.crown gall:根癌病 2.tomato canker:番茄细菌性溃疡病 3.fire blight(Erwinia amylovora):梨火疫病 4.Ralstonia solanacearum:青枯雷尔氏菌(番茄青枯病菌) 5.tent solarization:树体罩棚式日光热处理 6.ED80:80% 病原菌致死有效剂量 / 处理时长 |
9.4.5 对线生植物寄生线虫的防控效果
已有研究普遍证实:相较于防治植物病原真菌与田间杂草,土壤日光消毒在防控植物寄生线虫方面的效果稳定性更差(Lamberti、Greco,1991;McGovern、McSorley,1997)。研究提出假说:植物寄生线虫栖息土层更深,土壤太阳能消毒处理结束后,线虫能快速向上层土壤迁移,相较于病原真菌与杂草,线虫可更快再度侵染经太阳能消毒的地块,这也是太阳能消毒对线虫防效偏低的原因(Porter & Merriman,1983;Heald & Robinson,1987;Abdel-Rahim 等人,1988;Cartia 等人,1989;Stapleton & Heald,1991)。连续 2~3 年开展土壤太阳能消毒,能够延缓线虫对处理田块的再度定植(Candido 等人,2008);但 Sharma 与 Nene(1990)的试验表明,单次处理和连续两季田间太阳能消毒,对线虫种群的抑制效果相差不大。此外,受太阳能消毒处理后土壤内生物与非生物环境发生改变,该处理对土壤线虫种群密度的抑制效果有时不会立刻显现,反之部分抑制作用却可维持数月之久。斯塔普莱顿与德韦(1983)研究发现,太阳能土壤消毒对双螺旋盘旋线虫的防控效果在处理完成三个月后才逐步体现;而沃克和瓦赫特尔(1988)的试验表明,爪哇根结线虫幼虫被穿刺巴斯德杆菌侵染的概率,在土壤太阳能消毒后的十个月里持续上升。
自卡坦等人于 1976 年首次证实太阳能土壤消毒能够有效防治植物寄生线虫以来,已有大量研究围绕日光加温的杀线功效,针对多个属的植物寄生线虫开展试验探究。
关键专业注解 1.solarization(土壤太阳能消毒 / 日光土壤热处理):田间覆膜依靠太阳能升温焖土的绿色土壤消毒技术,无化学药剂,常用于土传病虫草防控; 2.phytopathogenic nematodes:植物病原线虫(植物寄生线虫); 3.recolonization:再度定植、土壤再侵染(有害生物从深层土回迁至耕作层并重建种群); 4.single- / two-season solarization:单季、连续两季田间太阳能土壤处理。 专有名词注释 1.soil solarization:土壤太阳能消毒(农田覆膜日晒高温灭病虫技术,业内通用译法) 2.Helicotylenchus digonicus Perry:双螺旋盘旋线虫(植物寄生线虫学名) 3.Meloidogyne javanica Treub:爪哇根结线虫 4.Pasteuria penetrans Sayre and Star:穿刺巴斯德杆菌(专性寄生根结线虫的生防细菌) 5.plant-parasitic nematodes:植物寄生线虫 |
波特与梅里曼(1983)研究证实,在 38~55℃温度区间内,外环线虫、爪哇根结线虫、穿刺短体线虫与半穿刺线虫对土壤太阳能消毒处理均高度敏感。多项豇豆田间试验结果表明,太阳能消毒可大幅压低肾形线虫虫口基数(希尔德、托马斯,1983;希尔德、罗宾逊,1987),但另有试验观测到该线虫在处理后会快速重返土壤定植(麦克索利、帕拉多,1986;夏尔马、内内,1990)。研究表明,覆盖地膜的厚度(而非地膜颜色)以及处理时节,会影响热处理对肾形线虫的抑制效果(科茨 - 贝克福德等人,1997、1998)。
土壤太阳能消毒对洋葱茎线虫防控效果优异:在意大利南部试验中,经过 4 周、6 周、8 周太阳能覆膜处理后,田间存活的洋葱茎线虫种群占比分别仅剩 10%、6% 和 2%(格雷科等人,1985);以色列受洋葱茎线虫重度侵染的蒜田,经太阳能消毒后,整个生长季大蒜均可正常收获(西蒂等人,1982)。
已有文献证实太阳能消毒能够高效防治胞囊类线虫:与空白对照土壤相比,经太阳能处理后的土壤在 5 厘米、10 厘米、15 厘米土层处,马铃薯金线虫卵孵化率分别下降 100%、68%、59%(拉蒙迪亚、布罗迪,1984)。意大利南部田间试验显示,太阳能消毒可显著抑制土壤及马铃薯根系上马铃薯金线虫的存活与繁殖(格雷科等人,2000),同时使胡萝卜胞囊线虫虫卵存活率降至 24%~38%(格雷科等人,1985)。此外,该处理对土壤及鹰嘴豆根系中的鹰嘴豆胞囊线虫(迪维托等人,1991)、豇豆胞囊线虫也具备明显的种群抑制作用(乔汉等人,1988;夏尔马、内内,1990)。
线虫学名中文定名(植保行业通用译名) 1.Mesocriconema xenoplax:外环线虫 2.Meloidogyne javanica:爪哇根结线虫 3.Pratylenchus penetrans:穿刺短体线虫 4.Tylenchulus semipenetrans:半穿刺线虫(柑橘根线虫) 5.Rotylenchulus reniformis:肾形线虫 6.Ditylenchus dipsaci:洋葱茎线虫(鳞球茎茎线虫) 7.Globodera rostochiensis:马铃薯金线虫 8.Heterodera carotae:胡萝卜胞囊线虫 9.Heterodera ciceri:鹰嘴豆胞囊线虫 10.Heterodera cajani:豇豆胞囊线虫 |
多项田间试验研究表明,采用土壤太阳能消毒技术防治根结线虫(根结线虫属Meloidogyne spp.)的效果并不稳定,部分试验出现防控失效的情况(格雷科等人,1985;巴伯切克、冯・布罗姆布森,1986)。多数研究认为,根结线虫卵块外部包裹的胶质基质可帮助虫卵抵御各类胁迫环境,是造成防效不佳的关键原因(多尔顿、努斯鲍姆,1961;奥利昂,1995);但尼科等人(2005)得出相反结论,该胶质基质并不会提升南方根结线虫虫卵在经太阳能消毒土壤中的存活能力。与此同时,也有大量田间试验证实太阳能热处理可高效防控根结线虫:意大利一项为期 3 年的试验显示,土壤太阳能消毒能显著减轻南方根结线虫对生菜、厚皮甜瓜的根部结瘤危害(兰贝蒂等人,2000);克罗地亚一项连续 4 年的试验发现,该技术可将土层深度 20 厘米范围内的根结线虫几乎完全灭杀(奥斯特雷茨、格鲁比希奇,2003);美国佛罗里达州的试验同样证实,土壤太阳能消毒能够降低南方根结线虫的田间虫口密度与根部受害程度(麦戈文等人,2002)。
温室环境下,土壤太阳能消毒对根结线虫通常能实现优异的防治效果(塞尼斯,1984;卡尔蒂亚等人,1989),不过塞浦路斯相关试验中,经 8 周土壤太阳能处理后,番茄田南方根结线虫侵染率仅下降 50%(约安努,2000)。近些年开展的温室试验进一步证明,太阳能消毒处理可大幅降低甚至近乎彻底根除根结线虫造成的植株侵染、根系结瘤问题,同时削减土壤内线虫种群数量(奥斯特雷茨、格鲁比希奇,2003;坎迪多等人,2008)(图 9.4)。
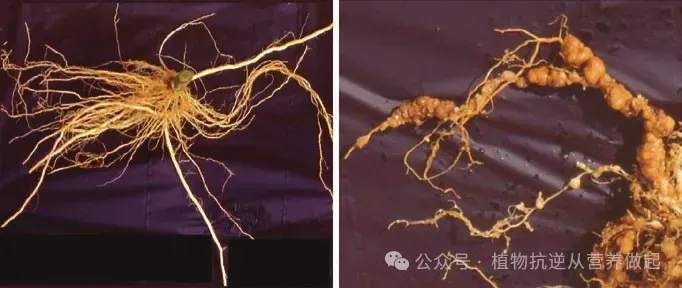
图 9.4 意大利南部塑料大棚内,土壤太阳能消毒对受南方根结线虫侵染番茄根系结瘤的影响
左侧:经太阳能消毒土壤培育的番茄根系;右侧:未做消毒处理土壤中的番茄根系,因大量根瘤畸形膨大。
土壤太阳能消毒对多种植物寄生线虫同样具备良好防效,涵盖索恩短体线虫(卡坦等人,1976;格林斯坦等人,1979b;格雷科等人,1990a)、香蕉穿孔线虫(巴塔查里亚、拉奥,1984)、尖尾潜根线虫(西瓦库马尔、马里穆图,1987)、盘旋线虫属线虫(夏尔马、内内,1990;科茨 - 贝克福德等人,1997、1998),以及危害水稻、小麦的滑刃线虫属、盘旋线虫属、潜根线虫属、短体线虫属、矮化线虫属线虫(斯特普尔顿、赫尔德,1991;波卡雷尔,1995;甘古利等人,1996)。克卢普费尔等人(2002)在桃园试验中发现,土壤太阳能消毒能够优化土壤微生物群落结构,富集可抑制外生根结线虫的有益微生物种群。美国佛罗里达州的田间试验表明,土壤经太阳能处理后,整个作物生长季内长尾刺线虫、环线虫属、异头矛线线虫的种群均受到显著抑制,但对细小拟毛刺线虫的防控效果不稳定(谢莱米等人,1993;麦克索利、麦戈文,2000;麦戈文等人,2002)。贝略等人(2004)提出,葡萄园重茬种植时土壤太阳能消毒难以有效防控葡萄传毒线虫葡萄剑线虫,原因在于热处理后葡萄根系仍可存活一年以上,线虫能依托残存根系继续繁衍。研究人员还在受限容积土壤中验证了太阳能消毒防控植物寄生线虫的应用潜力(吉布林 - 戴维斯、韦卡德,1988):黑聚乙烯套管封装土壤热处理可使柑橘半穿刺线虫、伤残短体线虫、外生根结线虫虫口数量下降 89%~100%(斯特普尔顿等人,1999);对土堆进行太阳能暴晒处理,可抑制 95% 的南方根结线虫卵孵化(尼科等人,2003)。育苗基质经太阳能消毒后,南方根结线虫与肾形线虫的虫口密度和侵染能力均显著下降,其中肾形线虫对高温更为敏感(高尔、丁格拉,1991)。
大量研究探究了太阳能消毒对土壤线虫群落整体的影响,一致表明该处理在灭杀植物寄生线虫的同时,也会大幅削减自由生活线虫的种群数量(斯特普尔顿、德韦,1983;斯特普尔顿、赫尔德,1991)。但相较于植物寄生线虫,自由生活线虫更易耐受高温处理,或是在消毒结束后快速重新定植土壤(斯特普尔顿,2000;奥斯特雷茨、格鲁比希奇,2003);纳斯尔・伊斯法哈尼(2007)研究证实,热处理配施有机肥,能够加快土壤线虫群落的恢复进程。奥弗曼(1985)对比试验显示,在降低土壤线虫总数量方面,土壤太阳能消毒效果优于绿肥覆田或除草剂休闲模式;卡尔曼等人(2006)也观测到,经太阳能消毒处理的稻田土壤线虫群落数量显著下降。针对不同营养类群线虫的专项分析表明:试验周期结束时,太阳能消毒对线虫群落结构造成的扰动基本消失;杂食性线虫耐热性最弱,其次是食细菌线虫与食真菌线虫,植食性线虫耐高温能力最强(王等人,2006)。
关于太阳能消毒对线虫的长效防控效果,现有试验结论差异明显:卡塔尔温室试验中,土壤太阳能消毒处理后一年内,黄瓜上不再出现根结线虫侵染的表观病症(阿塔 - 阿里,2007);但塞浦路斯与意大利南部的同类试验显示,处理满一年后,番茄田间的南方根结线虫、爪哇根结线虫侵染程度仅有小幅下降,甚至无明显变化(约安努,2000;坎迪多等人,2008)。
大田种植条件下,多项试验证实土壤太阳能消毒可连续两年显著减轻南方根结线虫对甘蓝、甘薯的危害(史蒂文斯等人,2003)。热处理后线虫快速回迁定植土壤,是太阳能消毒持效期偏短的核心诱因;而连年重复开展太阳能消毒,可延缓线虫再度侵染,逐步将虫口密度压低至防治经济阈值以下,持续强化杀线效果(坎迪多等人,2008)。
9.4.6 对杂草的影响
无论是大田试验还是温室试验中,利用 ** 土壤太阳能消毒(日光土壤覆膜热处理)** 开展杂草防控的相关研究十分广泛,但试验结果差异较大(埃尔莫尔,1991b;亚杜拉朱、米什拉,2004);在气候炎热的国家,该技术总能取得最优防除效果(阿尔 - 马苏姆等人,1993;萨希尔,1997)(图 9.5)。
学界推测,太阳能消毒调控杂草种群是多重作用机制共同作用的结果:高温改变杂草种子细胞代谢与细胞超微结构(辛格拉等人,1997);亚致死高温损伤种子后,土壤寄生微生物侵染劣变种子;覆膜升温打破杂草种子休眠;地膜密闭高温灼伤杂草地上茎叶(埃格利,1990;卡坦、德韦,1991)。除此之外,覆膜密闭环境下氧气、二氧化碳气体失衡,或是乙醛、乙烯等挥发性有毒物质释放,同样会造成杂草死亡(鲁宾、本杰明,1984;加姆利埃尔等人,2000)。
不同杂草品种对太阳能消毒处理的耐受能力差异显著(埃科诺穆等人,1997;埃尔莫尔,1998)。雷斯图西亚等人(1994)依据杂草耐热特性,将杂草划分为敏感型、耐受型、特性尚不明确三类(表 9.1)。其中敏感类杂草包含约 80 种一年生杂草,再细分为秋冬型萌发杂草、春夏型萌发杂草、无固定萌发时段杂草。相较于春夏季一年生杂草,越冬一年生杂草多在短日照、低温环境下萌发,耐热性更差,土壤小幅升温即可实现高效防除。仅需一周太阳能覆膜处理,就能有效防控早熟禾、春蓼、抱茎春美草、欧洲千里光等多数越冬敏感杂草(卡坦、德韦,1991);而夏季一年生杂草需要更高的覆膜地温或更长的热处理周期才能被杀灭(埃格利,1990)。耐太阳能消毒的杂草类群既包含一年生豆科(刺黄芪、蝎爪豆、蝎叶小冠花、槽荚草木犀)、菊科(加拿大蓬)、锦葵科(克里特花葵、尼斯锦葵)杂草,也包含香附子等多年生莎草类杂草。
马齿苋、苍耳、黄果龙葵等一年生杂草,以及狗牙根、石茅(约翰逊草)等多年生杂草被归为反应特性不明确类杂草。坎迪多等人 2006 年针对生菜田开展的太阳能消毒试验补充:田间种植条件下,大翅山黧豆(豆科杂草)属于耐太阳能消毒品种;而田蓟(多年生杂草)则对太阳能热处理表现敏感。
大量试验证实太阳能消毒对一年生杂草防效突出:斯塔普莱顿等人(2005)开展多年定点试验发现,日光覆膜升温处理可近乎 100% 灭杀多种一年生杂草,包含黄花草木犀、繁缕、一年生早熟禾、荠菜、马唐、斑地锦。另有多项大田与温室试验表明,苋菜、藜、臭荠、马唐、稗、牛筋草、牛膝菊、阿拉伯苜蓿、龙葵、苦苣菜等春夏、越冬型杂草的出苗受到土壤太阳能消毒近乎完全的抑制(埃尔莫尔,1993;莫亚、古川,2000;帕特里西奥等人,2006;坎迪多等人,2008)。利用太阳能消毒分别处理胡萝卜、茄子、番茄田,可高效防除埃及列当、皱叶列当;在高温季节开展覆膜处理时,两类列当的防控效果最优(雅各布索恩等人,1980;阿卜杜勒 - 拉希姆等人,1988;绍尔伯恩等人,1989)。多项番茄大田与温室试验显示,大麻列当、弯管列当在经过太阳能消毒的地块完全绝迹(阿布 - 伊尔迈莱,1991a;毛罗米卡莱等人,2005)。
现有文献普遍记载马齿苋耐太阳能消毒:有研究表明,种子需要 60℃处理 2 小时或 65℃处理 1 小时才会明显降低发芽率(韦杜、马斯,2004);但达尔奎斯特等人(2007)提出 39℃即可对马齿苋种子形成亚致死损伤,帕特里西奥等人(2006)的大田试验也证实,土壤太阳能消毒可大幅降低田间马齿苋发生量。关于菟丝子,阿布 - 伊尔迈莱、塔哈比(1997)认为其耐受太阳能消毒,而海达尔、伊斯坎德拉尼(1999)试验发现,太阳能处理能大幅消耗土壤中菟丝子种子库存。

图 9.5 意大利南部田间试验中土壤太阳能消毒对田间杂草发生的影响:左侧为太阳能消毒地块,甜瓜行间无杂草;右侧未做消毒处理,田间杂草疯长。
(正文页码:241 第 9 章 土壤太阳能消毒与可持续农业)
表 9.1 文献依据:依据对土壤太阳能消毒的敏感 / 抗性划分杂草种类
敏感型杂草
一年生秋冬生长型
蓝花琉璃繁缕、意大利疆南星、野燕麦、不育燕麦、玻璃苣、黑芥、荠菜、小荠、伊比利亚矢车菊、臭荠、茼蒿、金黄胡萝卜、刺酸模、牻牛儿苗属、香天芥菜、狐尾大麦、苦苣菜、宝盖草、多型苜蓿、一年生山靛、抱茎春美草、叙利亚苦刺、虞美人、佛氏虞美人、短穗虉草、奇异虉草、早熟禾、木贼蓼、野萝卜、春千里光、欧洲千里光、野芥菜、大蒜芥属、龙葵、苦苣菜、繁缕、细茎繁缕、膜质荨麻、小荨麻、婆婆纳属
一年生春夏生长型
苘麻、白苋、铺地苋、反枝苋、叙利亚红花、灰绿藜、小藜、墙生藜、鸭跖草、小蓬草、麦秆菊、曼陀罗、马唐、稗、水稗、牛筋草、大穗画眉草、犹太烟堇、墙烟堇、药用烟堇、软毛牻牛儿苗、裂叶番薯、小花锦葵、欧锦葵、埃及列当、皱叶列当、大麻列当、青狗尾草、绿狗尾、桃叶蓼、刺果黄花稔、龙葵、蒺藜、大巢菜、宾州苍耳、刺苍耳
多年生
盖氏虎尾草、田蓟(根茎繁殖)、药旋花、田旋花(种子 + 根茎繁殖)、狗牙根(种子繁殖)、问荆、节节草、酢浆草、车前属、石茅(种子繁殖)
抗性杂草
一年生
加拿大蓬、蝎叶小冠花、聚花牛舌草、刺黄芪、槽荚草木犀、蝎爪豆、克里特花葵、尼斯锦葵、大翅山黧豆
多年生
油莎豆、香附子
特性未明确杂草
一年生
苍耳、马齿苋、黄果龙葵
多年生
狗牙根(根茎繁殖)、石茅(根茎繁殖)
注:上标 a 数据引自坎迪多等,2006、2008
(页码 242 T达达博等)
专业名词注释(便于农学阅读) 1.Soil solarization:土壤太阳能消毒 / 日光覆膜土壤热处理(农田环保土壤消毒技术,透明地膜密闭覆膜,依靠日光升温土壤杀灭病虫杂草) 2.Orobanche:列当(全寄生恶性农田杂草) 3.Sublethal temperatures:亚致死温度(未直接致死、但损伤种子生理活性的高温) 4.Seed bank:土壤种子库(留存于土层内的杂草种子群体) |
多年生杂草相较一年生杂草更难防除,原因大概率是杂草繁殖体藏匿在土层深处,无法接触致死温度(鲁宾、本杰明,1984)。仅在日光土壤消毒处理后的表层 3~4 厘米土层内,才能观察到约翰逊草与田旋花种子受热失活(斯坦迪弗等,1984;卡坦、德韦,1991)。鲁宾与本杰明(1984)研究证实狗牙根、约翰逊草的根茎对高温敏感;但多项田间试验表明,日光土壤消毒处理反而会刺激香附子块茎萌发(埃格利,1983;库马尔等,1993;埃尔莫尔等,1997;迈尔斯等,2002;罗伊等,2004)。研究发现,莎草属杂草出苗受日光消毒期间剧烈的昼夜温差诱导,同时也受聚乙烯地膜理化性质影响(迈尔斯等,1996;韦伯斯特,2005)。大量研究证实,日光土壤消毒对多年生杂草、尤其是莎草属杂草防效不佳(杜兰蒂、库奥科洛,1988;罗斯科普夫等,1999;斯台普尔顿等,2005;坎迪多等,2008);另有文献记载该技术仅能实现杂草部分铲除(埃雷拉、拉米雷斯,1996;卡姆拉、高尔,1998),揭除覆膜后香附子还会快速重新萌发(利拉 - 萨尔迪瓦尔等,2004)。也有不少研究表明,日光消毒单用或搭配低剂量除草剂,均可有效防控莎草及其他多年生杂草(穆肖博齐等,1998;马伦科、卢斯托萨,2000;奥佐雷斯 - 汉普顿等,2001;吉尔里斯等,2005)。延长覆膜消毒周期至 8~10 周,可提升对香附子与其他多年生杂草的防除效果(鲁宾、本杰明,1983;蔡斯等,1998);还有试验显示,为期 90 天的日光消毒能大幅压低含香附子在内的田间杂草种群基数(史蒂文斯等,1990a;里奇等,2000)。搭配保温型覆盖地膜延长日光消毒时长,可通过高温灼伤地上杂草茎叶,大幅提升莎草防效(蔡斯等,1998、1999a),同时延长除草持效期、弱化种子埋深带来的避热影响(蔡斯等,1999b)。多项大田与温室试验中,帕特森(1998)发现,相较于传统黑白不透光聚乙烯膜,透光型日光消毒地膜可显著抑制甚至完全阻断香附子出苗与生长。
不同杂草物种的耐热性存在差异,致使日光土壤消毒无法彻底清除田间全部杂草群落。一项食用豆类大田普查结果显示,日光消毒可灭杀 80% 田间杂草,但对球根类、种子耐热、根系深扎或依靠多年生营养器官繁殖的杂草无抑制作用,甚至会促进其萌发(林克,1994)。马伦科、卢斯托萨(2000)统计发现,田间约 40% 杂草对日光消毒不敏感;萨图尔等(1991)也提出,不同杂草耐受土壤高温的能力参差不齐。塔米埃蒂、瓦伦蒂诺(2000)试验表明,单子叶杂草受日光消毒的防控效果劣于双子叶杂草;而阿卜杜勒 - 拉希姆等(1988)则认为高温处理对多数杂草防效优异,仅对野燕麦、反枝苋、马齿苋、苍耳、锦葵等一年生杂草,以及丝路蓟、狗牙根、莎草属、约翰逊草等多年生杂草作用有限。
覆膜时长、土壤最高温度、种子活力、种子埋土深度是决定日光消毒除草效果的四大关键因素(阿布 - 伊尔迈莱,1991b;埃尔莫尔,1991a)。霍洛维茨等(1983)指出,杂草防除效果和土层温度超 45℃的累计天数正相关,热敏性杂草所需消毒时间短于耐热型杂草(斯坦迪弗等,1984)。阿罗拉、亚杜拉朱(1998)证实,除草效果随种子入土深度增加而下降,深层土壤中的杂草种子常规避高温胁迫存活(埃格利,1983;霍洛维茨等,1983;鲁宾、本杰明,1984)。斯坦迪弗等(1984)测定,香附子、稗草种子仅在表层 3~4 厘米土层内失活,牛筋草、鸭跖草致死土层分别为 0~5 厘米、0~11 厘米;与之类似,早熟禾种子在表层 5 厘米消毒土内存活率下降,土层越深存活概率越高(皮奇等,2001)。因此,日光消毒后翻动土壤会破坏除草效果,深层存活杂草种子随翻土上浮造成田间二次侵染(埃格利,1983;阿布 - 伊尔迈莱,1991a)。土壤与种子含水量直接影响杂草高温致死:干燥种子环境温度升至 120℃仍可保有活性,吸水饱和的种子 50℃条件下即可死亡(鲁宾、本杰明,1984);土壤含水率 19% 时杂草种子可耐受 70℃高温 3 天,含水率降至 2% 时耐受时长可达 7 天(埃格利,1990)。多数试验证实,土壤湿润条件下开展日光消毒,除草效果优于干土处理(霍洛维茨等,1983;阿罗拉、亚杜拉朱,1998);但萨尔塞斯布埃诺等(2003)提出,消毒前基质持续高湿会抑制杂草出苗,也有学者认为消毒前单次灌水与多次补水的处理效果基本持平(格林斯坦等,1979c;霍洛维茨等,1983)。灌水措施对日光消毒土中独脚金的种子萌发与活性无明显影响(奥斯曼等,1991)。地膜颜色同样左右日光消毒除草活性:黑膜与透明膜均能压低杂草基数,但黑色地膜除草效果与持效期均不及透明聚乙烯膜(霍洛维茨,1980;霍洛维茨等,1983;斯坦迪弗等,1984;坎皮利亚等,2000)。阿布 - 伊尔迈莱(1991b)补充,日光消毒结束后加覆黑色地膜,能够进一步巩固除草成效。
日光土壤消毒成本低廉、除草稳定,同样适用于育苗床(帕特尔等,1995;埃莱夫塞罗里诺斯、詹诺波利蒂斯,1999;库马尔、夏尔马,2005)、盆栽基质(斯台普尔顿等,2002)以及新建果园园地(阿布 - 伊尔迈莱,1994)。日光消毒的除草持效期远长于对线虫、多数病原真菌的防效:坎迪多等(2008)监测发现,温室经日光消毒后两年内,田间一年生杂草与部分多年生杂草持续大幅减少乃至绝迹,狗牙根的抑制效果维持时间更久;另有橄榄园试验记录,单次日光消毒可实现园地连续 3 年无杂草(洛佩斯 - 埃斯库德罗、布兰科 - 洛佩斯,2001);番茄 - 黄瓜轮作地块中,香附子、油莎豆受日光消毒的抑制作用可横跨四个种植季(吉尔里斯等,2005)。消毒后尽量减少土壤扰动,能够进一步延长除草持效周期(贝尔、埃尔莫尔,1983)。
9.4.7 对作物生长与产量的影响
大量研究证实,日光土壤消毒可促进作物生长、提升收成(斯台普尔顿、德韦,1984;戴维斯,1991;加米尔、卡坦,1991),促生效果同样体现在育苗幼苗与落叶果树苗木上(斯台普尔顿、德韦,1982;萨莱诺等,2000);设施栽培环境下增效尤为突出,增产提质效果可延续两个以上种植茬口(坎迪多等,2008,图 9.6)。但也有负面研究结论:本达维德 - 瓦尔等(1997)发现,在消毒土壤播种的胡萝卜、洋葱生长受阻;科索内尔等(1998)提出,玉米沙拉菜减产是由于高温处理抑制了菌根侵染根系。除增产之外,多篇文献表明日光消毒还能提早蔬菜成熟期、缩短作物生育周期(陈等,1991;斯台普尔顿、德韦,1995;西尼加利亚等,2001;帕特里西奥等,2006)。

图 9.6 意大利南部塑料大棚内,太阳能热处理土壤中的番茄长势提升。背景地块为经太阳能热处理的土壤,番茄植株长势旺盛;前景地块是未做热处理的土壤,番茄长势弱小
(文献页码 246,T达达博等人)
格伦茨维希等人(1993)研究表明,土壤太阳能促生效果源于多项植株生理变化:热处理土栽培作物光合效率、蛋白含量上升,组织发育提速,生长后期衰老进程延缓。格伦茨维希团队(2000)测得,热处理土壤培育的番茄体内赤霉素含量显著升高,且赤霉素浓度与叶片干重呈线性正相关,说明土壤受热处理后,植株内源激素平衡也发生了改变。
多数研究者认为,太阳能热处理带来的作物增产效应并非单纯依靠抑病作用,即便在无土传病原菌的洁净土壤中该促生效果依然存在(阿布埃尔 - 梅吉德等,1998);其增产是土壤与植株多重理化、生物变化共同作用的结果:一是土壤可溶性矿质养分、有机质矿化产物、内源生长调节物质含量提升(凯恩、卡坦,1980;斯台普尔顿等,1984;凯恩等,1991、2000;格伦茨维希等,2000);二是土壤生物活性增强,次要土传病原菌得到抑制(格伦茨维希等,1993;加姆利埃尔、斯台普尔顿,1995;季阿莫斯、弗拉维尔,1995;勒比昂等,1997)。
9.5 土壤太阳能热处理与病虫害综合防治
病虫害综合防治是可持续农业的核心技术理念,指整合多种防控手段,将病虫危害控制在经济损失阈值以内。根据种植模式与田间生产条件因地制宜选配防控方案,既能充分发挥各类防治措施的效用,又可减少农药滥用(马伦等,1997;佩兰,1997;马丁,2003)。土壤太阳能热处理适配绝大多数种植制度,可与化学药剂、生物防治、农艺栽培措施兼容甚至协同增效,在病虫害综合防控体系中应用潜力突出(斯台普尔顿、德维,1995;卡坦,2000;斯台普尔顿,2000)。随着综合防控理念普及、广谱土传病虫害治理需求提升,各国科研人员陆续开展试验,探究太阳能热处理搭配各类替代防控技术,协同治理土传真菌、线虫与杂草的作用机理与实际效果。
太阳能热处理配合减量熏蒸药剂,可进一步强化对多数病原真菌的抑制效果,但在长期连年熏蒸的地块,该协同增效作用不明显(阿尔布雷茨等,1996)(见图 9.7)。已有试验证实:太阳能热处理分别搭配溴甲烷、威百亩、1,3 - 二氯丙烯 + 氯化苦、棉隆,可协同降低大丽轮枝菌、尖孢镰刀菌、齐整小核菌的活体基数、发病概率与田间病症(弗兰克等,1986;本 - 耶普赫特等,1988;埃谢尔等,2000;于塞尔等,2007);同时有效管控丝核菌、仙人掌疫霉、辣椒疫霉(于塞尔,1995;本利奥格鲁等,2005)、烟草疫霉、尖孢镰刀菌、齐整小核菌(切莱米等,1994、1997;史蒂文斯等,2003;切莱米、米鲁索,2006)、番茄疫霉、陆生根腐疫霉(季阿莫斯,1984;波特等,1989)以及芸薹根肿菌(波特等,1991)。药剂与热处理的施用顺序直接影响防效:埃谢尔等人(2000)发现,田间先亚致死高温闷土、后施用熏蒸剂的防治效果,远优于先施药后热处理的操作顺序。此外,热处理前先用威百亩润湿土壤,还能额外防控多种常规难除土传病原物(弗兰克等,1986;乔斯沃尔德,2000)。
低剂量熏蒸型杀线虫剂(1,3 - 二氯丙烯、二溴乙烷、溴甲烷、威百亩、棉隆)与太阳能热处理联用,可显著提升土壤杀线虫能力(斯台普尔顿、德维,1983;巴伯切克、冯布伦布森,1986;卡尔蒂亚等,1989;于塞尔等,2007)。美国试验显示,减量 1,3 - 二氯丙烯搭配太阳能闷土,可大幅压低番茄、辣椒根结线虫与马铃薯马铃薯胞囊线虫种群数量(拉蒙迪亚等,1986;切莱米、米鲁索,2006);意大利田间试验中,该组合对胡萝卜胡萝卜胞囊线虫、洋葱茎线虫防控效果突出(格雷科等,1990、1992)。颗粒剂型杀线虫剂配合太阳能热处理,能有效减轻生菜、甜瓜受南方根结线虫侵染形成的根瘤(兰贝尔蒂等,2000);但美国加州部分田间试验表明,药剂联用热处理对南方根结线虫、桃根腐线虫无增效作用(斯台普尔顿等,1987)。
皮奇等人(2001)研究证实,低用量威百亩结合太阳能热处理,能够强化对杂草的灭杀效果。
(页码 247 第 9 章 土壤太阳能热处理与可持续农业)

图 9.7 意大利南部塑料大棚茄子栽培试验长势对比:前景小区为太阳能热处理地块,中间小区为未做热处理地块
(页码 248,T达达博等人)
外源有机物料抑病机理一般认为包含两点:一是有机质改良驱动土壤微生物群落向拮抗菌优势菌群转变;二是有机质腐解过程释放有毒抑菌物质(斯特林,1988)。有机物料与太阳能热处理存在协同效应,正是源于上述两条作用路径被进一步强化;加姆利埃尔、斯台普尔顿(1997)测定,有机物料腐解产生的大量挥发性抑菌物质,会在覆膜热处理的密闭土壤环境中大量富集。
大量研究证实,堆肥、作物残体、绿肥、畜禽粪污等各类有机培肥物料配施太阳能热处理,可高效防控多种土传病害(儿玉、福井,1982;弗里曼、卡坦,1988;加姆利埃尔、斯台普尔顿,1993a、b;切莱米等,1997)。高氮有机物料可大幅削减包括炭腐病菌这类耐热病原菌在内的各类土传病原物与线虫基数(春、洛克伍德,1985;洛达,1995;罗德里格斯 - 卡瓦纳,1986)。有机物料搭配太阳能热处理增效显著:有机质分解生成的氨、亚硝酸可在覆膜密闭土壤中留存更久、扩散更充分,抑菌效果大幅提升(德维、卡坦,1991;拉扎罗维茨等,2001;洛达等,2003)。鸡粪还田结合太阳能闷土,可有效抑制生菜终极腐霉(加姆利埃尔、斯台普尔顿,1993)、草莓丝核菌与仙人掌疫霉(本利奥格鲁等,2005)、番茄齐整小核菌(史蒂文斯等,2003);尿素或农家粪肥配施热处理,能显著降低孜然尖孢镰刀菌、菜豆炭腐病菌的田间侵染基数(洛达,1995)。恩迪亚耶等(2007)采用富氮谷子秸秆配施太阳能热处理,显著降低豇豆炭腐病菌接种量与发病程度;但屠宰废弃物搭配热处理无明显防效(库尔特、埃米尔,2004)。肉鸡垫料、棉籽饼、羽毛粉、豆粕、园林枯枝等有机废弃物与太阳能热处理复配,相较单一处理,对根结线虫的抑制能力全面提升(加姆利埃尔、斯台普尔顿,1993b;史蒂文斯等,2003;切莱米,2006;奥卡等,2007)。格雷科等人(1992)发现,土壤预先施用小麦秸秆再进行太阳能热处理,对洋葱茎线虫防效提升,但该组合无法防控胡萝卜胞囊线虫。黎巴嫩田间试验表明,鸡粪改良后覆膜热处理,可强力抑制列当、田野菟丝子种子萌发,实现杂草绿色防控(海达尔等,1999;海达尔、西达赫迈德,2000);马莱克等(2007)提出,在气温偏低、闷晒周期不足的不利条件下,洋葱、大蒜干残体还田可补强太阳能热处理的除草效果。
十字花科作物残体在土壤腐解过程中会释放异硫氰酸酯、醛类等有毒挥发性物质,已有研究证实这类挥发物能够抑制或降低多种土传病虫的种群基数(安格斯等人,1994;凯纳思,1996;梅顿等人,1996;马蒂森、柯克高,2006)。加姆利埃尔等人(2000)研究表明,地膜覆盖形成的局部厌氧环境与土壤温度,直接决定土壤中挥发性有毒物质的浓度。
早在 20 世纪 80 年代初,就有报道指出将十字花科绿肥与太阳能土壤消毒技术配施可提升土壤太阳能消毒效果(堀内等人,1982),此后数十年间,各国科研人员在多种试验条件下针对该配套技术开展了大量试验研究。室内试验发现,亚致死热处理配施十字花科植物残体,能够抑制终极腐霉与齐整小核菌的孢子萌发(斯泰普尔顿等人,1995);大田试验中,该组合处理可有效防治菜豆壳球孢(洛达等人,2003)。大田条件下,太阳能土壤消毒搭配十字花科还田物料,对甘蓝尖孢镰刀菌专化型、菜豆壳球孢防效突出(拉米雷斯 - 维拉普杜瓦、芒内克,1987、1988;索萨,1994;洛达,1995),同时还能强化对其余各类土传病原菌的抑制作用(加姆利埃尔、斯泰普尔顿,1993a、1993b;加姆利埃尔等人,2000)。太阳能消毒前翻压芥菜饼粕或新鲜植株残体,可大幅削减黄瓜尖孢镰刀菌专化型与菜豆壳球孢的病原接种体量(洛达等人,1997;洛达、马瓦尔,2000;伊斯雷尔等人,2005),但甘蓝残体还田无法提升太阳能消毒对疫霉菌土壤种群的抑制效果(科埃略等人,1999)。温室栽培模式下,太阳能消毒结合十字花科生物熏蒸技术,可有效防控番茄黑斑根腐病菌引发的栓根病(迪亚斯埃尔南德斯等人,2005),同时高效杀灭甜瓜、辣椒根结线虫与田间杂草(普洛格、斯泰普尔顿,2001;格雷罗等人,2005)。
非十字花科物料配施太阳能消毒同样具备增效作用。平克顿等人(2000)研究显示,太阳能消毒搭配苏丹草、大麦绿肥,可显著增强对樟疫霉、大丽轮枝菌的抑制能力;弗洛雷斯 - 莫克特苏马等人(2006)利用银胶菊绿肥结合太阳能热处理,减轻了小核菌对洋葱幼苗的危害。此外,利拉 - 萨尔迪瓦尔等人(2004)证实,三齿拉瑞阿叶片树脂浸出液与太阳能消毒联用,既能降低土传病害发病率,还可对线虫起到部分防控效果。布洛赫等人(2000)提出假说:这类非十字花科绿肥之所以能和太阳能消毒产生协同增效,核心机理同样是植株腐解释放有毒活性物质。
已有文献证实无机改良物料也能优化太阳能土壤消毒效果(斯泰普尔顿等人,1990)。氰氨化钙配施太阳能消毒,可近乎彻底铲除火龙果栽培土壤中的尖孢镰刀菌,以及黄瓜田的葫芦科腐皮镰刀菌专化型(布尔博斯等人,1997;崔等人,2007);麦克索利、麦戈文(2000)采用太阳能消毒配合碳酸氢铵或硫酸铵撒施,有效降低了长尾短体线虫在土壤中的种群密度。
太阳能消毒与生防菌剂联用,是优化土传病虫害治理的又一可行路径(卡坦,2000)。热处理后,生防真菌哈茨木霉不会出现种群衰减,反而能在作物根际逐步增殖(波拉斯等人,2007a;贾亚拉杰、拉达克里希南,2008)。切特等人(1982)在温室试验中发现,鸢尾种球裹覆哈茨木霉菌剂,可大幅减轻立枯丝核菌、齐整小核菌诱发的病害;大田与温室环境下,太阳能消毒配施哈茨木霉能显著防治番茄镰刀菌冠腐根腐病(于塞尔、奇纳尔,1989;西万、切特,1993)。土壤太阳能消毒结束后施用哈茨木霉,可使蜜环菌病原彻底丧失侵染活性(奥蒂耶诺等人,2003),显著压低番茄腐霉猝倒病发病程度(贾亚拉杰、拉达克里希南,2008)。但也存在例外:太阳能消毒搭配哈茨木霉,无法协同增效防控菜豆立枯丝核菌、黄瓜终极腐霉、罗勒尖孢镰刀菌专化型(米努托等人,1995)。
其余多种生防资源同样可以提升太阳能消毒土壤的抑病能力。绿色粘帚霉与太阳能消毒组合施用,可作为番茄、辣椒白绢病(齐整小核菌)的优选防控方案(里斯泰诺等人,1991、1996);荧光假单胞杆菌菌剂施入经太阳能消毒的土壤,能够减少番茄、凤仙花立枯病与腐霉病害发生(麦戈文等人,2002;贾亚拉杰、拉达克里希南,2008),降低生姜青枯病(青枯雷尔氏菌)发病率(阿尼思等人,2000)。商品化链霉菌制剂配施太阳能消毒,对腐霉、立枯丝核菌以及镰刀菌、轮枝菌枯萎病增效明显,但温室条件下防控番茄栓根病菌效果不稳定(麦戈文等人,2002;米努托等人,2006)。育苗圃中,太阳能消毒配合枯草芽孢杆菌灌根,可削减发根农杆菌诱发的樱桃砧木根瘤(古普塔、科斯拉,2007);太阳能消毒地块播种甜菜时,拌入枯草芽孢杆菌、哈茨木霉或镰刀菌复合拮抗菌种,能够缓解立枯丝核菌造成的危害(加索尼等人,2007)。生防制剂还能强化太阳能消毒对线虫的防效:太阳能消毒搭配促生根际细菌或坚强芽孢杆菌商品化菌剂,对番茄、辣椒根结线虫的防效与化学药剂持平(科卡利斯 - 布雷勒等人,2002;吉安纳库等人,2007)。太阳能消毒后施用穿刺巴斯德氏菌制剂,可协同抑制黄瓜上的爪哇根结线虫、南方根结线虫(措察扎基斯、高恩,1994);但若搭配淡紫拟青霉商品化药剂,无法进一步提升热处理的杀线效果(阿纳斯蒂阿迪斯等人,2008)。哈彻、梅兰德(2003)提出,太阳能消毒结合生防菌,可治理单独热处理难以杀灭的耐热型杂草。
优化农艺管理模式、完善田间土壤管控,同样能够放大太阳能消毒的应用成效。约安努(2001)选用嫁接在抗病番茄砧木上的茄子幼苗定植于太阳能消毒地块,实现了黄萎病、栓根腐病、根结线虫全防控;约翰逊等人(2007)采用夏季长时间太阳能消毒结合休闲翻耕,将香附子杂草基数压至经济危害阈值以内;索托马约尔等人(1999)联用太阳能消毒与土壤淹水,成功防控花生根结线虫。佩兰等人(1998)指出,合理维护菌根共生体系能够替代土壤化学熏蒸,加姆利埃尔等人(2004)也证实,丛枝菌根真菌与太阳能消毒协同作用,可显著抑制细香葱粉根病发生。
9.6 小结
现有文献对可持续农业给出了多种定义,但其核心要义趋于一致:在保障种植业经济效益的前提下,不造成环境污染、不损耗农田与自然资源,兼顾土壤、水体及生物多样性保护(多兰,2002;弗朗西斯等人,2006)。太阳能土壤消毒技术契合可持续农业发展理念,既能绿色高效治理各类土传有害生物,助力农产品实现无农药生产、提升产品市场溢价空间。
成本收益分析表明,依托更低的投入成本,太阳能消毒的投入产出比优于多数化学防控手段(亚龙等人,1991;埃尔莫尔,1991a;贝尔,1998;埃斯佩兰奇尼等人,2003;哈辛等人,2004);该技术可与绝大多数病虫害防治措施兼容配套,便于嵌入综合植保体系,是其另一大应用优势。
但气候条件、栽培制度的地域局限性制约了太阳能消毒大范围推广:该技术在温暖区域的温室园艺、露地蔬菜与果园夏茬应用时防效稳定,冷凉地区使用不仅防效下降、用药成本攀升,也不适用于大面积雨养大田粮食作物。
业内普遍提及的地膜残膜难回收、集约化种植模式下消毒周期偏长等短板,可通过地膜材料改良加以破解:选用可生物降解地膜解决残膜污染问题,高导热地膜或多技术联施方案则能缩短土壤暴晒消毒时长;复合处理还能改善深层土壤的受热效果,弥补常规太阳能消毒土层深处热量不足、无热效应的缺陷。
纯太阳能消毒后续应用重心大概率集中于温室栽培:温室作物经济附加值高,环保收益与经济效益双向拉动技术落地。依托同等逻辑,该技术在苗圃育苗基质、苗床预处理消毒领域潜力巨大(乔贝、达南杰,2003),也可用于温室、果园定植前的病原真菌与线虫消杀(詹森、布扎德,1988;斯泰普尔顿等人,1989;邓肯等人,1992;里格等人,2001)。大田灌溉农田遭遇病虫害无登记农药可用、药剂药害风险高或用药成本过高,或是单一热处理能够兼治多种病虫时,太阳能消毒是理想的绿色土壤处理方案(埃尔莫尔,1990);有机农场、临近居民区禁限用化学熏蒸药剂的地块,同样是太阳能消毒的优先应用场景。


 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
