仲恺农业工程学院:牛病毒性腹泻病毒感染诱导ROS-HIF-1a轴驱动的糖酵解重编程,其通过损害RIG-I依赖的I型干扰素应答来增加病毒复制
牛病毒性腹泻病毒(BVDV)属于瘟病毒属,是引发全球养牛业严重经济损失的重要病原体,其感染可导致肠炎、免疫抑制乃至持续性感染。BVDV的持续性感染严重依赖其精密的天然免疫逃逸策略——尤其是抑制I型干扰素介导的抗病毒应答——然而,病毒如何重塑宿主代谢以服务于这一目的,其背后的分子机制尚不为人熟知。
2026年5月7日,仲恺农业工程学院研究团队在Journal of Virology上发表题为“Pestivirus bovine viral diarrhea virus infection induces ROS–HIF-1a axis-driven glycolytic reprogramming, which increases viral replication by impairing RIG-I-dependent type I interferon response”的最新研究论文。研究结果揭示了BVDV利用ROS−HIF-1α−糖酵解轴减弱MAVS介导的抗病毒信号传导并促进病毒复制的新机制。
IF:4.17 中科院2区 | JCR/Q2 病毒学 参考译文:瘟病毒属牛病毒性腹泻病毒感染诱导ROS-HIF-1a轴驱动的糖酵解重编程,其通过损害RIG-I依赖的I型干扰素应答来增加病毒复制 第一作者:Yuan Li 通讯作者:Yigang Xu |
1.BVDV感染期间乳酸和IFN-Ⅰ水平的变化
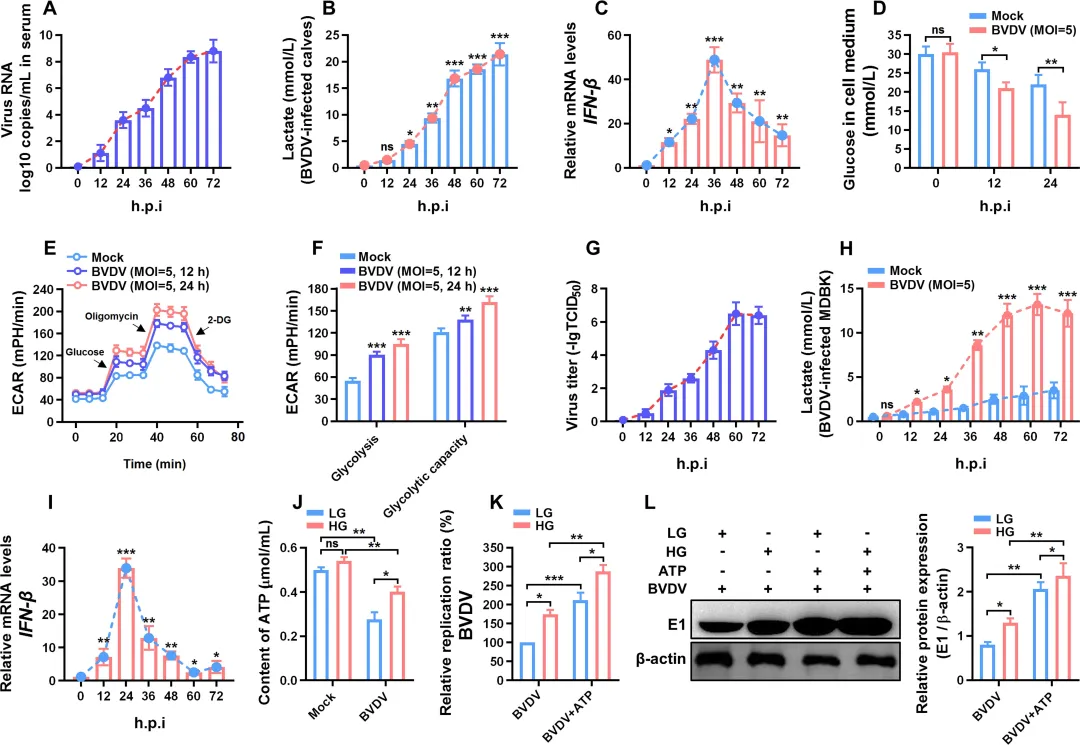
为探究BVDV感染后乳酸与I型干扰素水平的变化,研究者首先在感染BVDV的犊牛体内进行了动态检测。结果发现,感染后犊牛血液中的乳酸水平呈现持续性升高(图1B);相比之下,IFN‑β的mRNA表达在感染后约36 h达到峰值,随后显著下降(图1C),提示乳酸积累与IFN‑I抑制存在时间上的关联。进一步的体外实验使用牛肾细胞(MDBK)验证了这一现象:BVDV感染显著增强了细胞的葡萄糖消耗和细胞外酸化率(ECAR),表明糖酵解活性被激活(图1D–F)。与体内结果一致,感染MDBK细胞中乳酸产量随时间逐步升高(图1H),而IFN‑β mRNA在感染后24 h达到最高水平后迅速回落(图1I)。此外,在高糖培养条件下,细胞内ATP含量较高,且外源性ATP补充可明显促进BVDV的复制(图1J–L)。
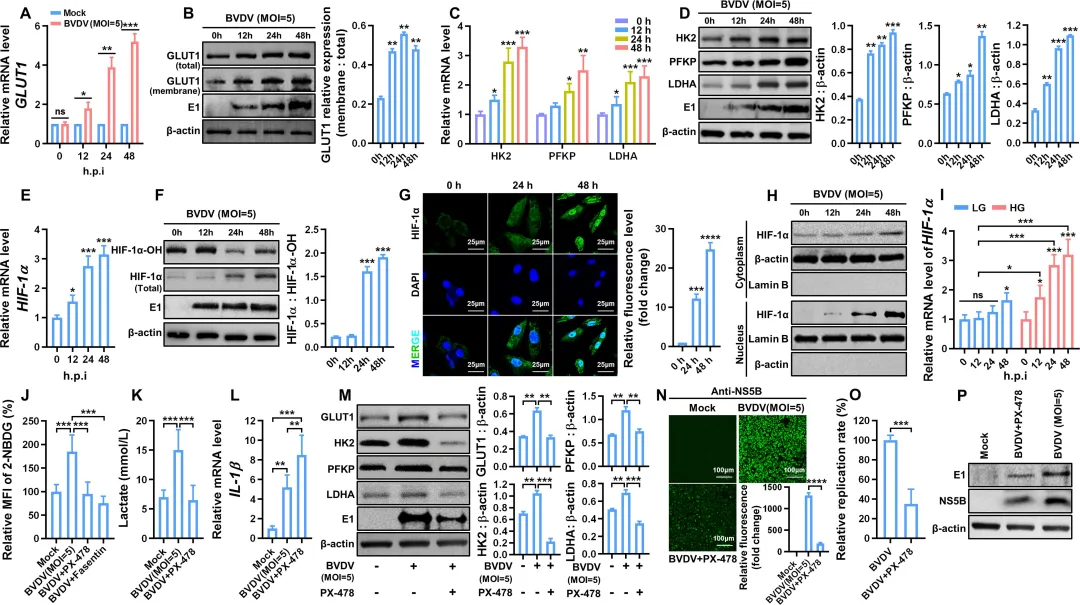
图1.BVDV感染后乳酸产量、细胞外酸化率(ECAR)、IFN-β表达水平和细胞内ATP含量的测定2.BVDV感染通过增强HIF-1α的表达和稳定性促进糖酵解研究者检测了BVDV感染后糖酵解关键蛋白的表达变化。结果显示,感染MDBK细胞中葡萄糖转运蛋白GLUT1的mRNA水平随时间显著上调,且GLUT1向细胞膜的转位明显增强(图2A–B)。同时,糖酵解关键酶HK2、PFKP和LDHA在mRNA及蛋白水平均呈现时间依赖性升高(图2C–D)。鉴于HIF-1α是上述基因的核心转录调控因子,进一步分析发现:BVDV感染不仅上调HIF-1α mRNA和蛋白表达,还显著降低其羟基化水平(HIF-1α‑OH),提示HIF-1α稳定性增加(图2E–F)。免疫荧光和核质分离实验证实,HIF-1α的核转位在感染后明显增强(图2G–H)。使用HIF-1α抑制剂PX-478或GLUT1抑制剂Fasentin预处理后,BVDV诱导的葡萄糖摄取和乳酸生成显著减少,而IFN‑β表达则明显回升(图2J–L)。此外,PX-478可下调GLUT1、HK2、PFKP和LDHA的蛋白水平,并显著抑制病毒复制(图2M–P)。图2.BVDV感染增强了HIF-1α的稳定性和表达,以促进感染MDBK细胞中的糖酵解3.BVDV感染通过ER应激-ROS轴促进HIF-1α的表达和稳定性
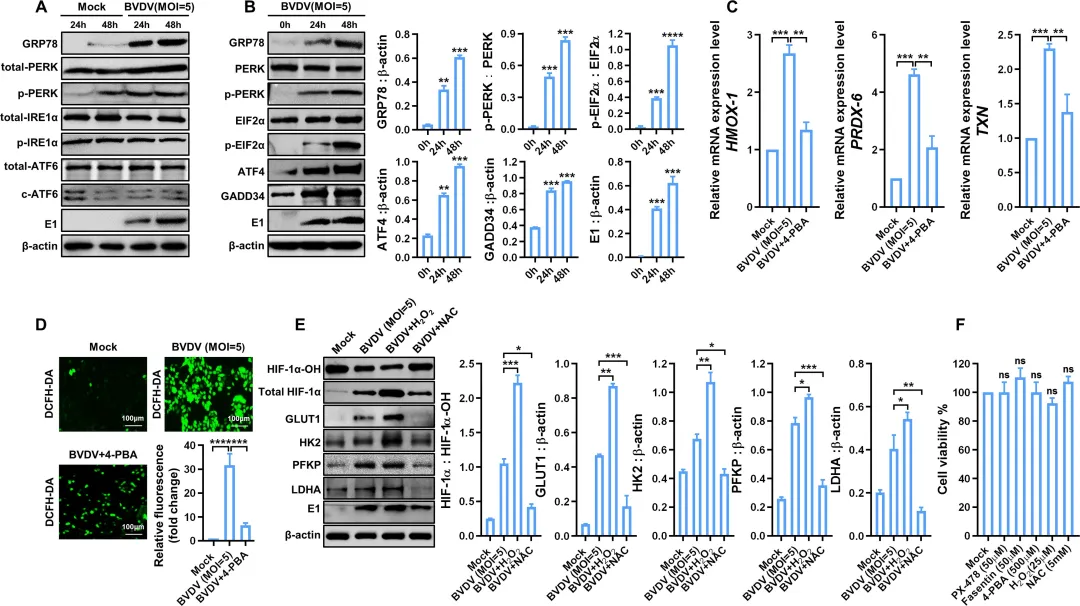
为阐明BVDV上调HIF-1α的上游机制,研究者检测了内质网应激通路的变化。结果显示,BVDV感染后GRP78蛋白表达显著升高,且仅PERK‑eIF2α分支被激活,而IRE1α和ATF6通路无明显变化(图3A–B)。使用ER应激抑制剂4‑PBA处理后,氧化应激相关基因HMOX‑1、TXN和PRDX‑6的表达被显著抑制(图3C),同时BVDV诱导的活性氧水平也明显下降(图3D)。进一步探究ROS对HIF-1α的作用:以H₂O₂(ROS诱导剂)处理可降低羟基化HIF-1α水平,上调GLUT1、HK2、PFKP和LDHA蛋白表达;相反,ROS抑制剂NAC则升高羟基化HIF-1α水平,并抑制这些糖酵解蛋白的表达(图3E)。细胞活性检测显示所用抑制剂和激活剂在实验浓度下无明显细胞毒性(图3F)。图3.BVDV感染通过ER应激-ROS轴促进HIF-1α的表达和稳定性4.BVDV通过糖酵解抑制RLR信号通路
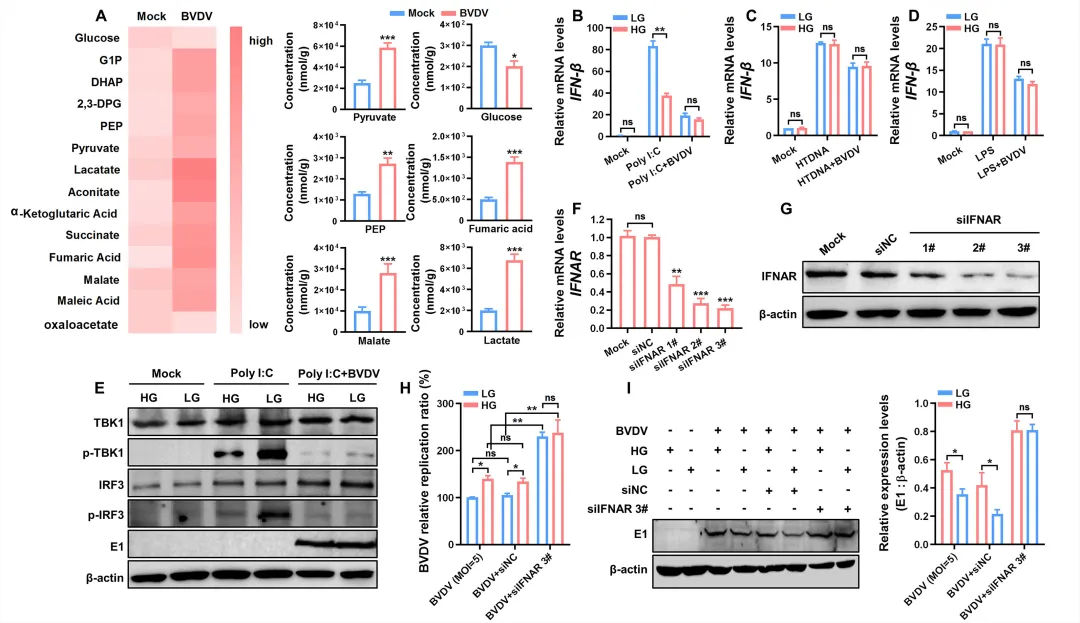
为明确BVDV诱导的糖酵解具体抑制哪条天然免疫通路,研究者首先检测了感染后细胞的糖代谢产物。结果显示,多种糖酵解中间产物(如丙酮酸、乳酸、DHAP等)显著升高,而草酰乙酸水平下降,确认糖酵解通量增强(图4A)。随后分别使用cGAS‑STING通路激动剂HT‑DNA、TLR通路激动剂LPS和RLR通路激动剂poly(I:C)处理细胞。结果发现,与高糖培养组相比,低糖条件下poly(I:C)诱导的IFN‑β mRNA表达显著增强(图4B),而HT‑DNA或LPS处理组之间无明显差异(图4C–D)。进一步分析表明,高糖环境下poly(I:C)诱导的TBK1和IRF3磷酸化水平明显低于低糖环境(图4E)。为验证糖酵解通过抑制IFN‑I产生而促进病毒复制,研究者敲低IFNAR后,低糖培养不再能限制BVDV复制,病毒RNA和蛋白表达在高、低糖条件下无显著差异(图4F–I)。5.BVDV感染促进HK 2-MAVS-VDAC 1复合物形成,以抑制RLR-MAVS途径激活
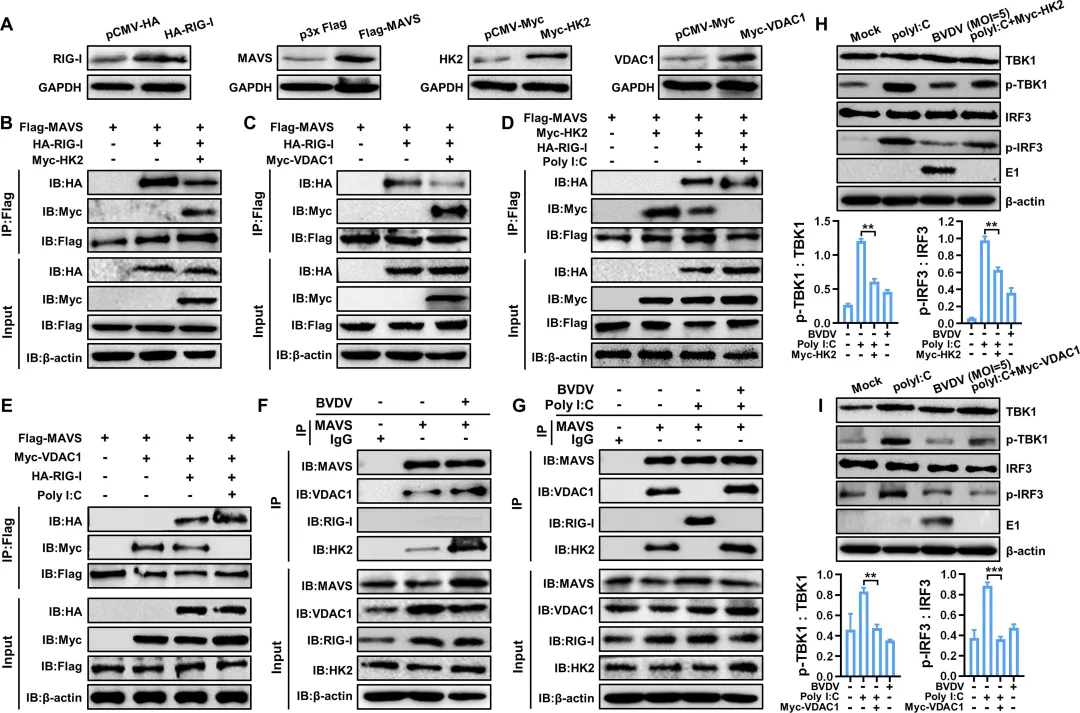
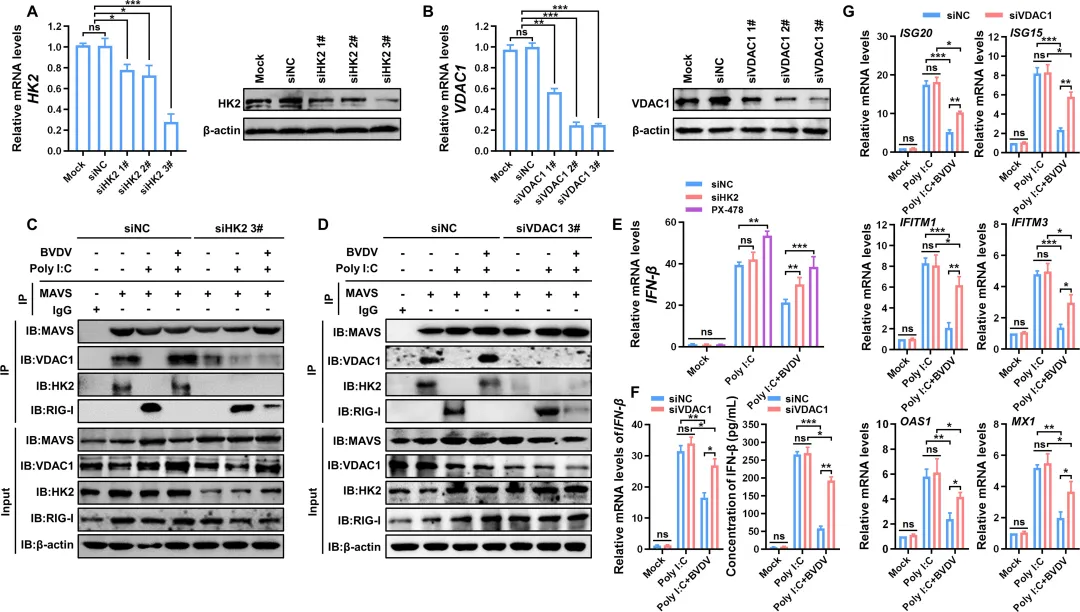
为阐明BVDV诱导的糖酵解如何抑制RLR信号通路,研究者首先通过免疫共沉淀证实,HK2和VDAC1分别能与MAVS相互作用,共同形成HK2‑MAVS‑VDAC1三聚体复合物(图5B–C)。该复合物的形成会竞争性阻断RIG‑I与MAVS的结合(图5D–E)。在BVDV感染的细胞中,内源性HK2‑MAVS及VDAC1‑MAVS的相互作用显著增强,而poly(I:C)诱导的内源性RIG‑I‑MAVS结合则被明显抑制(图5F–G)。功能上,过表达HK2或VDAC1均能降低TBK1和IRF3的磷酸化水平(图5H–I)。进一步通过siRNA敲低HK2或VDAC1,发现内源性RIG‑I‑MAVS的相互作用得以恢复(图6C–D),同时IFN‑β的mRNA表达和蛋白分泌均显著上调(图6E–F)。此外,敲低VDAC1还可增强多种干扰素刺激基因(如ISG20、ISG15、IFITM1等)的表达(图6G)。图5.BVDV感染促进HK 2-MAVS-VDAC 1复合物的形成,以抑制RLR-MAVS途径活化图6.抑制HK 2-MAVS-VDAC 1复合物形成可增强RLR信号通路介导的IFN-β产生6.糖酵解促进BVDV复制
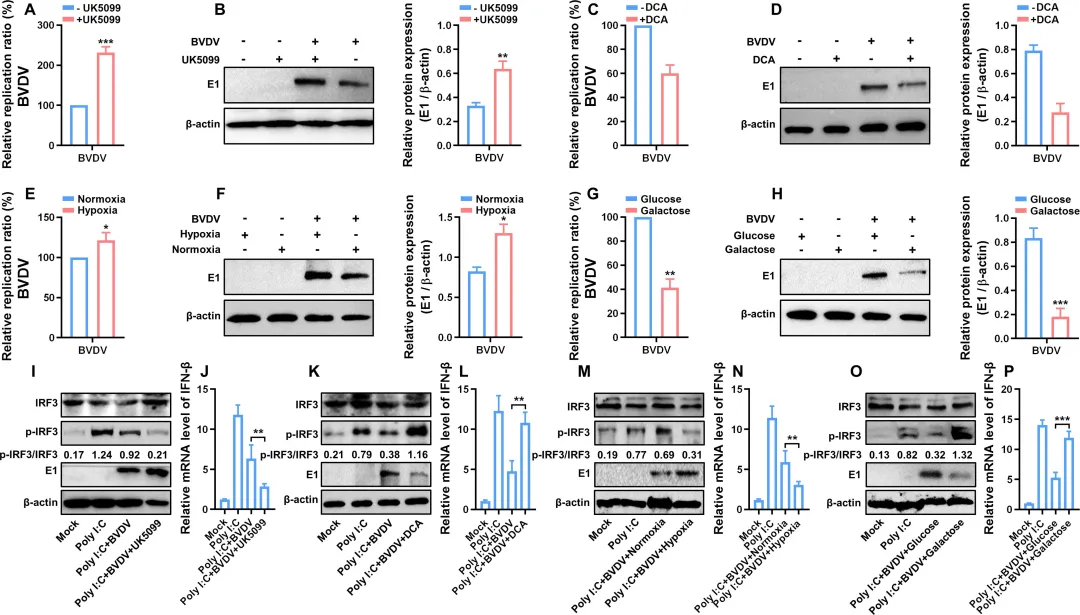
为验证BVDV诱导的糖酵解是否直接促进病毒复制,研究者采用多种代谢干预手段。结果显示,抑制丙酮酸线粒体转运(UK5099处理)显著增强病毒RNA复制和蛋白表达(图7A–B);而激活丙酮酸氧化(DCA处理)则强烈抑制病毒复制(图7C–D)。缺氧条件(5% O₂)下,糖酵解进一步增强,病毒复制明显高于常氧组(图7E–F);将培养液中的葡萄糖替换为半乳糖(迫使细胞转向氧化磷酸化)后,病毒复制显著下降(图7G–H)。进一步分析机制发现:UK5099处理或缺氧均降低了IRF3的磷酸化水平及IFN‑β表达(图7I–J、M–N),而DCA处理或半乳糖培养则恢复IRF3磷酸化并上调IFN‑β(图7K–L、O–P)。7.糖酵解介导的乳酸产生通过减弱IFN-1应答促进BVDV复制
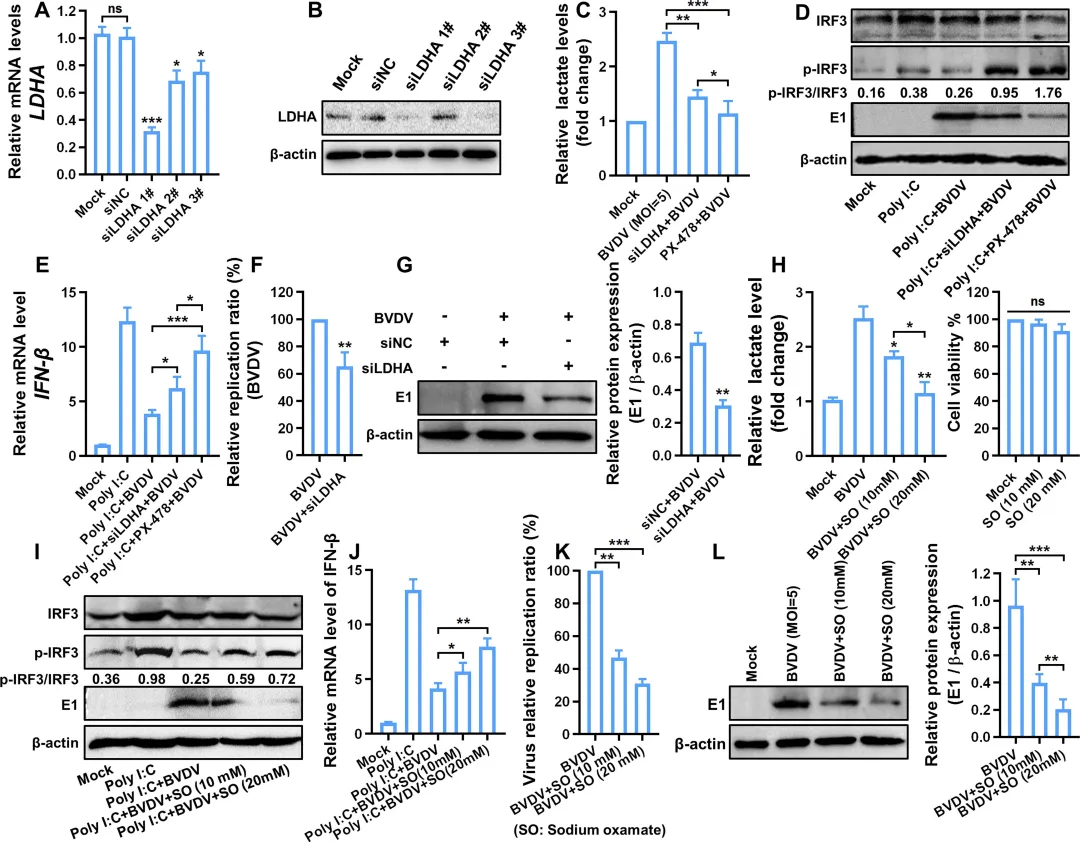
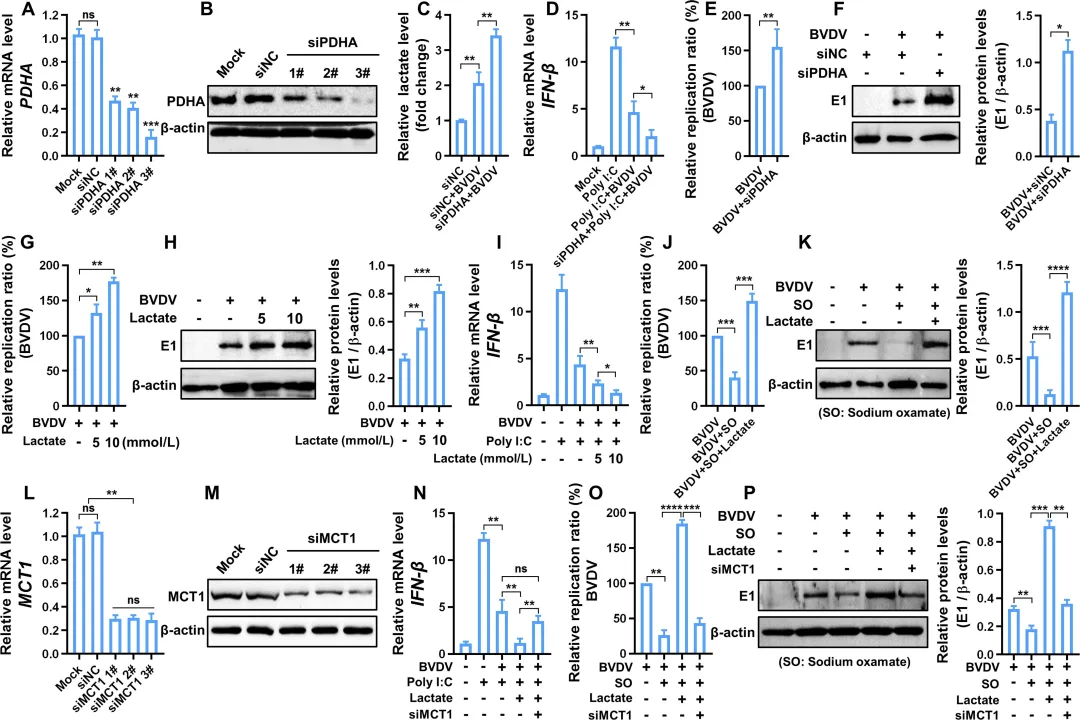
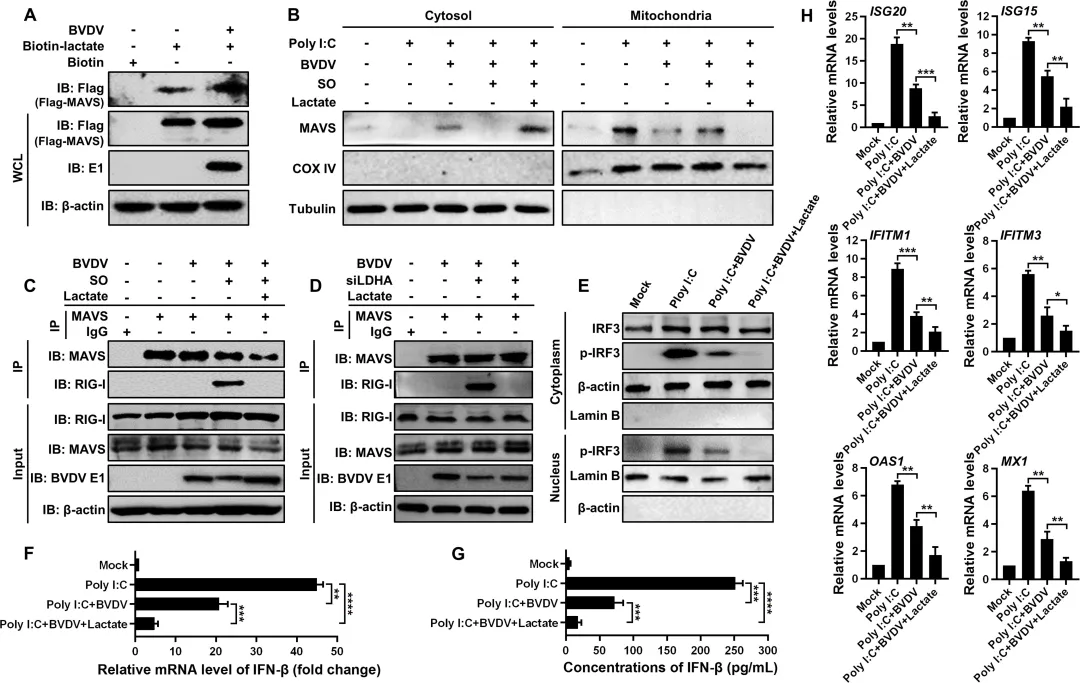
为探究乳酸在BVDV诱导的糖酵解促复制效应中的作用,研究者首先敲低乳酸生成关键酶LDHA,发现乳酸产量显著下降(图8C),同时IRF3磷酸化水平升高,IFN‑β表达增强(图8D–E),病毒RNA和蛋白水平均被明显抑制(图8F–G)。使用LDHA抑制剂草酸钠也得到一致结果:乳酸减少伴随IRF3磷酸化降低和IFN‑β表达上升,病毒复制呈剂量依赖性受抑(图8H–L)。相反,敲低丙酮酸脱氢酶PDHA(迫使丙酮酸转向乳酸生成)导致乳酸积累、IFN‑β表达下降,病毒复制增强(图9A–F)。外源性补充乳酸可直接抑制IRF3磷酸化及IFN‑β产生,并促进病毒复制(图9G–K)。机制上,生物素标记乳酸pull‑down实验显示乳酸直接结合MAVS(图10A),并促使MAVS从线粒体转位至胞质(图10B),从而破坏RIG‑I‑MAVS相互作用(图10C–D),导致IRF3核转位受阻(图10E),最终抑制IFN‑β及多种ISGs的表达(图10F–H)。此外,敲低乳酸转运蛋白MCT1可阻断外源乳酸的促病毒效应(图9L–P)。图8.糖酵解产生的乳酸盐通过减弱IFN-I应答增强BVDV复制8.MCT 1依赖性乳酸转运减弱IFN-Ⅰ应答,促进BVDV感染
为明确乳酸跨膜转运在BVDV感染中的作用,研究者靶向单羧酸转运蛋白MCT1进行干预。首先设计并验证了靶向MCT1的siRNA,其敲低效率经qRT‑PCR和Western blot确认(图9L–M)。在poly(I:C)预处理的BVDV感染细胞中,外源性补充乳酸可显著抑制IFN‑β的表达并促进病毒复制;然而,敲低MCT1后,外源乳酸的细胞内摄取受到明显阻碍,导致IFN‑β mRNA表达水平较对照组显著回升(图9N)。与此一致,MCT1敲低还强烈抑制了BVDV的RNA复制和病毒蛋白表达(图9O–P)。图9.抑制PDHA和乳酸转运蛋白MCT 1可减弱IFN-1应答,从而促进病毒感染9.乳酸通过直接结合MAVS负责糖酵解介导的RLR信号传导抑制
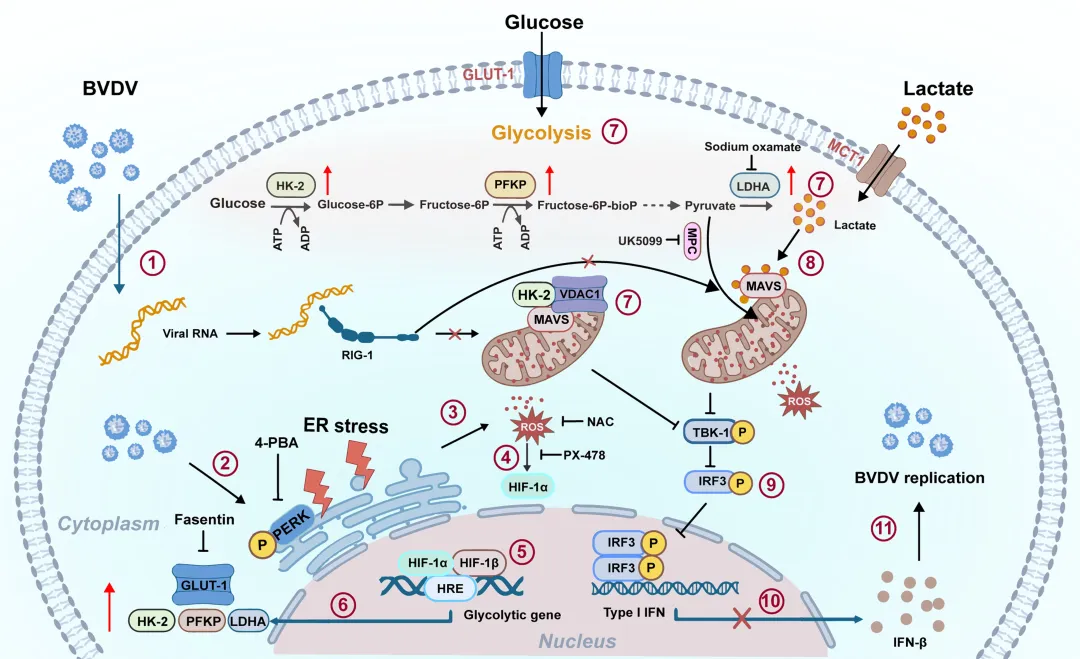
为阐明乳酸抑制RLR信号通路的直接分子机制,研究者首先通过生物素标记乳酸pull‑down实验证实,乳酸可直接与MAVS蛋白结合(图10A)。进一步亚细胞定位分析发现,乳酸处理可诱导MAVS从线粒体转位至细胞质,而抑制糖酵解来源乳酸产生的草酸钠则能阻断BVDV诱导的MAVS胞质转位(图10B)。功能上,使用草酸钠或敲低LDHA均可显著增强BVDV感染细胞中内源性RIG‑I与MAVS的相互作用;而外源性补充乳酸则能逆转这一增强效应,恢复对RIG‑I‑MAVS结合的抑制(图10C–D)。乳酸介导的RIG‑I‑MAVS复合物破坏进一步导致IRF3磷酸化水平下降及其核转位受阻(图10E)。功能检测结果显示,外源乳酸补充可显著降低IFN‑β的mRNA表达和蛋白分泌,并抑制多种干扰素刺激基因(包括ISG20、ISG15、IFITM1、IFITM3、OAS1和MX1)的转录(图10F–H)。图10.乳酸通过直接结合MAVS抑制RLR信号通路本研究首次揭示了BVDV通过激活内质网应激-活性氧轴,稳定并上调HIF-1α表达,从而驱动糖酵解重编程。感染后HK2、LDHA等关键酵解酶表达显著升高,乳酸大量积累,为病毒复制提供能量支持的同时,也为后续免疫逃逸奠定代谢基础。
机制上,BVDV诱导的糖酵解通过两条平行途径抑制RIG‑I/MAVS介导的I型干扰素应答:一方面,HK2与MAVS及VDAC1形成三聚体复合物,物理阻断RIG‑I与MAVS的结合;另一方面,乳酸直接结合MAVS并促使其从线粒体转位至细胞质,进一步破坏信号复合物的形成。这种“双重打击”策略高效削弱了宿主抗病毒免疫。
源论文链接:https://journals.asm.org/journal/jvi on 13 May 2026 by 58.58.135.38
编辑:九九
投稿合作:shouyiyanquan@163.com
可为课题组代发文章宣传、新闻通稿、招聘等
| 标注“原创”仅代表原创编译,原文版权归原作者所有。本文仅用作学术交流分享,如有错误或侵权请后台私信订正或删除。 |