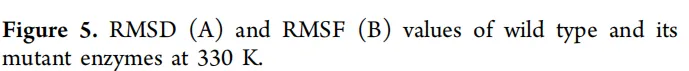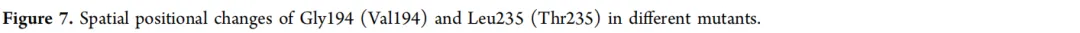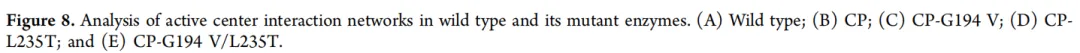2026 年 3 月 9 日,来自南京农业大学的Huibing Chi等人在Journal of Agricultural and Food Chemistry上发表了一篇题为Overcoming the Stability−Activity Trade-Off in Lipoxygenase by Integrated Computational-Assisted Structure-Guided Design的研究性论文。脂氧合酶(LOX)在食品加工和制药研究中的工业潜力常常受到酶活性与热稳定性之间持续存在的权衡限制。在此,应用集成计算辅助的结构导向设计,在集胞藻(Nostoc sphaeroides)LOX(NsLOX)中构建了一个突变体(CP-G194V/L235T),该突变体有效地解除了这一限制。与野生型相比,该工程化变体在50°C下的半衰期显著延长了45.29倍(达到389.08 ± 36.62分钟),熔融温度(Tm)为67.85 ± 0.40°C,对亚油酸的比活力提高了2.89倍(达到215881.25 ± 5024.47 U/mg)。结构和分子动力学分析表明,这些突变通过增强疏水相互作用和优化表面静电作用来增强结构刚性,同时诱导α-螺旋发生细微重排,从而扩大了底物结合腔并改善了底物通道的可及性。本研究证明了已建立的半理性设计工具在克服LOX工程化中的一个关键限制方面的有效应用,并且这是首次报道在LOX酶中同时成功提高热稳定性和比活力。
Introduction
脂氧合酶(LOX,EC 1.13.1.13)属于氧化还原酶类,其结构特征为含有非血红素铁。它特异性催化含有顺,顺-1,4-戊二烯结构的多不饱和脂肪酸的加氧反应,生成具有共轭双键的脂肪酸氢过氧化物。因此,LOX可添加到面粉中以提高白度、增加面筋含量、改善面团水合作用并提升面制品品质。此外,LOX在制备天然风味化合物、生产医药产品、纸浆漂白以及增强卷烟香气等方面具有广泛应用。目前,商品化LOX产品主要从大豆中提取。然而,大豆中多种同工酶的存在导致提取的LOX产品批间稳定性差,阻碍了质量控制和大规模应用。微生物来源的LOX具有不依赖原材料来源、成本效益高、易于大规模生产等优势,已成为研究热点。然而,已报道的微生物来源LOX通常存在热稳定性差和活性低的问题,增加了微生物污染的风险和整体生产成本,从而对工业应用提出了挑战。因此,通过蛋白质工程探索具有更高稳定性和活性的LOX至关重要。
增强蛋白质热稳定性的一条成熟策略是强化其结构中的柔性区域[8,9]。与具有较高柔韧性的嗜冷蛋白相比,嗜热蛋白通常以其整体刚性增加而闻名。一个显著特征是蛋白质中连接表面更结构化α-螺旋和β-折叠的不规则环区。与相对稳定、在皮秒到纳秒时间尺度上发生局部微小波动的α-螺旋和β-折叠不同,环区通常在微秒到毫秒范围内表现出更大振幅的运动,并发挥着重要的功能作用。由于其更大的柔韧性,环区在热胁迫下更容易发生导致蛋白质去折叠的构象变化。研究人员通过降低枯草芽孢杆菌168 L-天冬酰胺酶N端一个高柔性环区的柔韧性,成功提高了其催化效率和热稳定性。类似地,通过调整B因子和吉布斯去折叠自由能,并结合多序列比较,改变了黑曲霉CBS513.88甘露聚糖酶的柔韧性,开发出了热稳定性显著提高的突变体。因此,目前提高热稳定性的创新方法涉及针对蛋白质内热敏感柔性区域的“热点”进行分子修饰。
分子动力学模拟被广泛用于研究蛋白质的柔韧性。该方法通过模拟蛋白质在不同温度下如何改变形状,并观察蛋白质分子在原子水平上随时间推移的运动情况,研究人员可以预测蛋白质的哪些区域最可能首先去折叠。分子动力学中使用的一个关键指标是均方根波动值,它衡量蛋白质结构的稳定性。较高的RMSF值表明蛋白质的特定部分振动更大,在模拟过程中比其平均位置更具柔韧性。值得注意的是,蛋白质的热稳定性和活性之间往往存在平衡。通过增加蛋白质刚性来提高热稳定性有时会导致其活性下降。因此,寻找稳定蛋白质而不损害其催化活性的方法仍然是一个挑战。
大量研究工作致力于解决这一限制。例如,采用涉及四轮定向进化的分子进化策略来克服葡萄糖氧化酶的稳定性-活性权衡。类似地,基于共有序列的半理性设计与组合突变相结合已应用于缓解超氧化物歧化酶的这种权衡。一种整合了进化分析、共有序列设计和二硫键工程的多策略工程化方法成功解决了木聚糖酶的稳定性-活性权衡问题。此外,Luo等人开发了一种整合能量与进化探索、共有序列分析和二硫键策略的方法,以解决纳豆激酶的稳定性-活性权衡。尽管取得了这些进展,但在解决脂氧合酶稳定性-活性权衡方面的进展有限。此外,阐明此类酶中权衡分子基础的机制研究仍然很少。
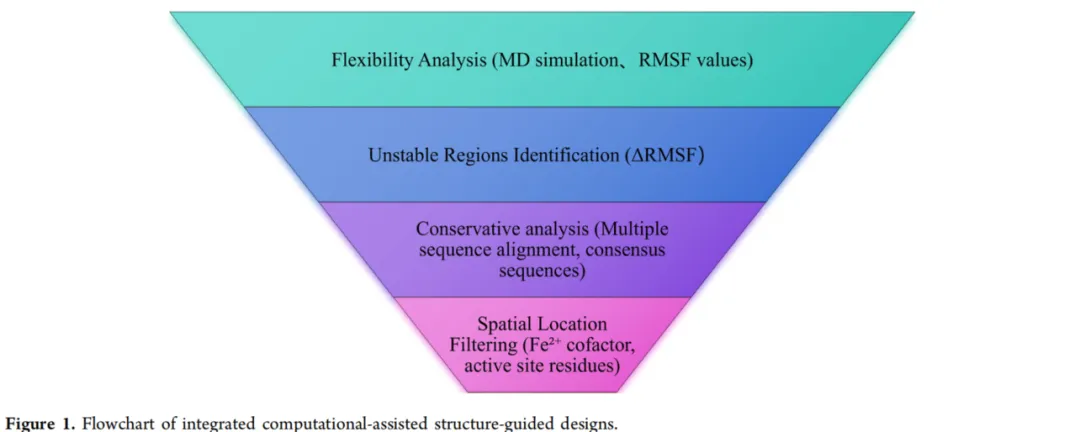
为了弥补这一空白,应用了一种结合分子动力学模拟、多序列比对、结构分析和保守性分析的集成计算策略,以克服脂氧合酶的稳定性-活性权衡。虽然先前已通过B因子引导的分子修饰提高了集胞藻LOX(NsLOX)的热稳定性,获得了热稳定性显著提高的双突变体T2C/S36P(CP),但为了工业应用仍需进一步优化。通过这种综合计算方法和两轮组合诱变,获得了一个成功解耦稳定性-活性权衡的组合突变体。随后的序列疏水性和柔韧性分析、分子动力学模拟以及三维结构建模揭示了性能提升的分子机制。这些发现不仅为其他LOX的分子修饰提供了见解,也为该领域的未来研究提供了有价值的视角。
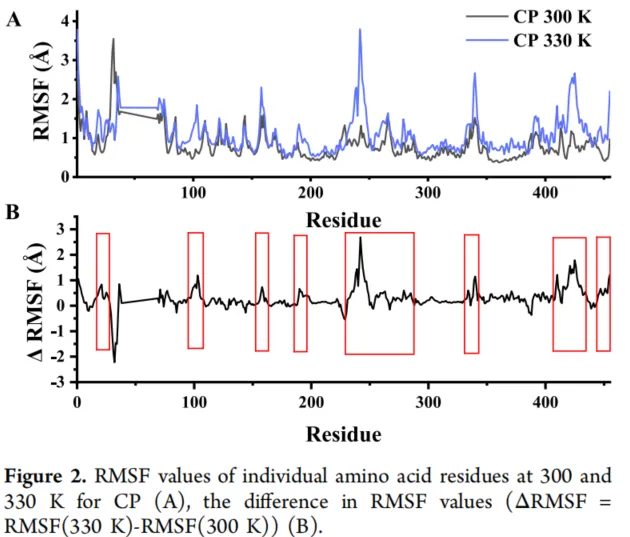
过度的柔韧性会损害蛋白质的稳定性,而增强柔性区域的刚性已被证明是提高蛋白质稳定性的有效策略。RMSF值较高的区域波动幅度较大,表明不稳定性增加。为了研究突变酶CP对温度变化特别敏感的区域,分别在300 K(26.85°C)和330 K(56.85°C)下进行了分子动力学模拟(图2A)。残基40−68的RMSF呈现线性趋势,因为该区域在初始晶体结构中缺乏电子密度,因此其结构未被解析,在模拟开始时是缺失的。通过分析330 K和300 K之间RMSF值的差异(ΔRMSF)来识别柔性区域。值得注意的是,在以下区域观察到显著的ΔRMSF值:Tyr17−Lys28、Arg96−Leu105、His157−Glu159、Val190−Leu198、Asn231−Pro283、Ala332−Leu341、Ser412−Arg437和Ala445−Ile455,表明这些区域在响应高温时柔韧性增加(图2B)。因此,这八个区域被确定为NsLOX中的热稳定区域。
为了进一步确定控制CP热稳定性的关键残基,应用了保守性和空间位置分析。首先,与来自BRENDA数据库的热稳定LOX同源物进行多序列比对,识别出进化保守性低的柔性区域。从这些区域中,排除了符合以下任一标准的残基:(1)物理化学性质高度相似的残基(例如,Ile/Leu、Val/Leu、Asp/Glu);(2)保守模式不适合诱变的位点;(3)先前已表征的Thr243位点,已知其稳定性提升效果甚微。此外,需要强调的是,保守氨基酸对蛋白质功能至关重要,突变可能导致活性丧失。共有序列分析确定了四个对酶功能至关重要的绝对保守残基(Leu337、Ile418、Arg437、Met451),为保持催化完整性,这些残基被排除。通过这种基于保守性的筛选,得到了11个候选残基:Gly194、Leu235、Pro249、Ile336、Val357、Phe365、Ser368、Trp369、His372、Lys373和Ser412。
随后的空间位置分析进一步精简了该列表。空间过滤排除了靠近功能元件的残基:Fe²⁺辅因子10 Å以内的残基(Ser368、Trp369、His372、Lys373)以及活性位点残基5 Å以内的残基(Phe365、Ser412)。通过这种集成的保守性和结构分析,确定了五个关键残基(Gly194、Leu235、Pro249、Ile336和Val357)作为NsLOX热稳定性的主要决定因素。选择这些位点进行后续的饱和突变,以最大化产生改良变体的概率。
通过测定野生型和突变体酶在50°C热处理后的残余活性,确定了关键氨基酸残基对NsLOX热稳定性的影响。与野生型和突变酶CP相比,突变酶CP-G194V、CP-G194A、CP-L235T、CP-P249D和CP-L336V的热稳定性显著提高。值得注意的是,突变酶CP-L235T、CP-G194V和CP-G194A在热处理后残余活性最高,分别达到95.05 ± 1.96%、93.04 ± 2.19%和90.37 ± 0.98%,而突变酶CP-P249C和CP-A426P的残余活性仅为79.81 ± 2.22%和82.47 ± 2.62%。
为了进一步提高NsLOX的热稳定性,选择了CP-G194V和CP-L235T的组合。该策略基于功能互补原则:G194V显著提高酶活性,而L235T显著提高热稳定性(表2)。同时,结构分析显示Gly194和Leu235位于催化口袋的两个相邻α-螺旋上(图7),表明它们之间存在潜在的空间协同调控作用。因此,设计此组合旨在探索可能的协同效应,并积极验证突变之间的上位相互作用。在构建过程中,以CP-L235T为模板,通过定点突变在Gly194位点引入Val突变,成功获得了组合突变体CP-G194V/L235T。热处理后,该突变酶的残余活性测定为96.12 ± 1.74%,显著高于野生型和CP突变体(p < 0.05)(表1)。
经Ni²⁺亲和层析纯化后,突变酶CP、CP-G194V、CP-L235T和CP-G194V/L235T以及野生型酶均表现出高纯度和相似水平的蛋白表达量。为了评估蛋白质的热力学稳定性,测定了突变酶的半衰期和熔融温度(Tm)值(表2)。突变体CP-G194V、CP-L235T和CP-G194V/L235T在50°C下的半衰期显著延长,分别为286.55 ± 11.97分钟、345.31 ± 4.81分钟和389.08 ± 36.62分钟。这些数值对应为野生型酶(8.59 ± 0.12分钟)的33.36倍、40.20倍和45.29倍。一级失活曲线与这些半衰期趋势一致。类似地,CP-G194V、CP-L235T和CP-G194V/L235T的Tm值分别为66.25 ± 0.34°C、67.01 ± 0.32°C和67.85 ± 0.40°C,均显著高于野生型(59.81 ± 0.17°C),与CP(65.34 ± 1.74°C)相比分别提高了0.91°C、1.67°C和2.51°C。此外,突变酶CP-G194V的比活力为215881.25 ± 5024.47 U/mg,显著高于野生型(78080.00 ± 1732.87 U/mg)和突变酶CP(197832.16 ± 3860.89 U/mg),比野生型提高了2.76倍。CP-L235T的比活力为65472.18 ± 5525.32 U/mg,略低于野生型酶。然而,该突变并未对组合突变酶CP-G194V/L235T的比活力产生负面影响;相反,它协同地将突变体的比活力提高至225613.6 ± 11944.25 U/mg,是野生型的2.89倍。总之,所有突变酶的热稳定性均显著增强,且CP-G194V和CP-G194V/L235T突变体的比活力显著提高。此外,与野生型和单点突变体相比,双突变体CP-G194V/L235T表现出显著的协同效应。其催化效率不仅显著高于野生型,也明显高于两个亲本单点突变体,表明G194V和L235T突变在增强活性方面产生了正协同作用。同时,该双突变体的热稳定性与最稳定的单点突变体L235T相当,成功实现了将高活性和高稳定性整合于单一变体的设计目标。这些结果证实了基于功能互补和结构邻近性的组合设计策略的有效性,并揭示了G194V和L235T突变位点表现出显著的正功能协同性。
此外,为了研究所观察到的协同效应是否具有底物混杂性,进一步测定了该酶及其突变体对第二偏好底物——α-亚麻酸(ALA)的催化活性。结果发现,所有酶变体对ALA的比活力均低于对亚油酸(LA)的比活力,表明关键突变并未改变酶固有的底物偏好顺序。具体而言,野生型NsLOX对ALA的活性测定为64294.25 ± 1876.43 IU/mg。在突变体中,CP和CP-G194V的活性分别提高至109528.88 ± 2592.33 IU/mg和130268.77 ± 5214.06 IU/mg,而CP-L235T和CP-G194V/L235T的活性分别为31711.63 ± 4731.32 IU/mg和58325.68 ± 1804.89 IU/mg。这些结果表明,某些突变(CP和CP-G194V)对催化活性的增强作用在一定程度上可延伸至底物ALA。
为了进一步评估最优突变体CP-G194V/L235T的稳定性,研究了其在工业条件下的化学耐受性。突变体CP-G194V/L235T在各种化学胁迫条件下均表现出优异的稳定性。在含有10% DMSO的缓冲液中,突变体的残余活性与空白对照组无显著差异,表明其对有机溶剂DMSO具有高耐受性。相比之下,在含有0.1% Tween-20的缓冲液中,尽管其稳定性较对照组略有下降,但仍显著优于野生型酶:突变体的半衰期维持在小时级别,而野生型酶在相同条件下的半衰期仅为8.95分钟。这一比较清楚地表明,通过理性设计增强的结构稳定性能够有效抵御工业催化中常见的化学胁迫,为突变体CP-G194V/L235T的实际应用提供了关键的实验支持。
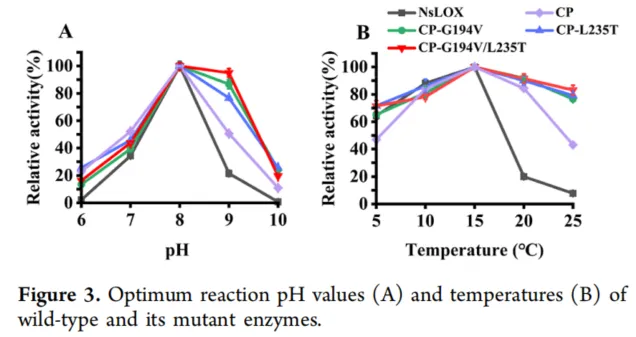
为了更深入地研究突变如何影响NsLOX的其他酶学性质,测定了突变酶CP-G194V、CP-L235T和CP-G194V/L235T的最适反应pH值和温度。如图3A所示,突变酶和野生型NsLOX的最适反应pH均为8.0。值得注意的是,突变酶的pH范围比野生型酶更宽;突变酶CP-L235T和CP-G194V/L235T在pH 9时相对活性保持在80%以上,而在pH 6.0和10.0时保持在20%左右。相比之下,野生型酶在这些pH水平下的催化活性显著降低,凸显了突变酶优异的pH稳定性。如图3B所示,突变体和野生型NsLOX的最适反应温度均为15°C。有趣的是,在15°C以上,突变酶CP-G194V、CP-L235T和CP-G194V/L235T表现出比野生型酶更高的相对催化活性。具体而言,它们在20°C和25°C时的相对活性分别高于90%和80%,超过了野生型酶和CP-L235T。这表明突变酶的热稳定性显著增强。
动力学参数在评估酶结合底物和催化反应速率的能力方面起着至关重要的作用。以LA为底物测定了突变酶的动力学参数,结果见表3。突变酶CP-G194V和CP-G194V/L235T的Km值分别为5.14 ± 0.78 μM和15.99 ± 1.24 μM,均显著低于野生型(19.46 ± 1.24 μM)。此外,它们的kcat/Km值从野生型的472.85 μM⁻¹·s⁻¹分别提高到CP-G194V的750.24 μM⁻¹·s⁻¹和CP-G194V/L235T的638.63 μM⁻¹·s⁻¹。突变酶对底物亚油酸的亲和力和催化效率显著增强(p < 0.05)。尽管突变酶CP-L235T的Km值为19.03 ± 0.61 mM,与野生型相比无显著变化;但其kcat/Km为427.73 μM⁻¹·s⁻¹,低于野生型。这表明该突变酶的催化效率降低,与比活力降低的实验观察结果一致。然而,突变酶CP-L235T并未影响CP-G194V/L235T的催化性能。
4.1.1. 野生型及其突变体酶的二级结构含量分析为了研究突变如何影响NsLOX的二级结构特征,对野生型及突变体酶进行了远紫外圆二色光谱分析。随后根据野生型和突变体酶的圆二色光谱确定了二级结构含量。结果表明,与野生型相比,所有突变体酶的二级结构均未发生显著改变。
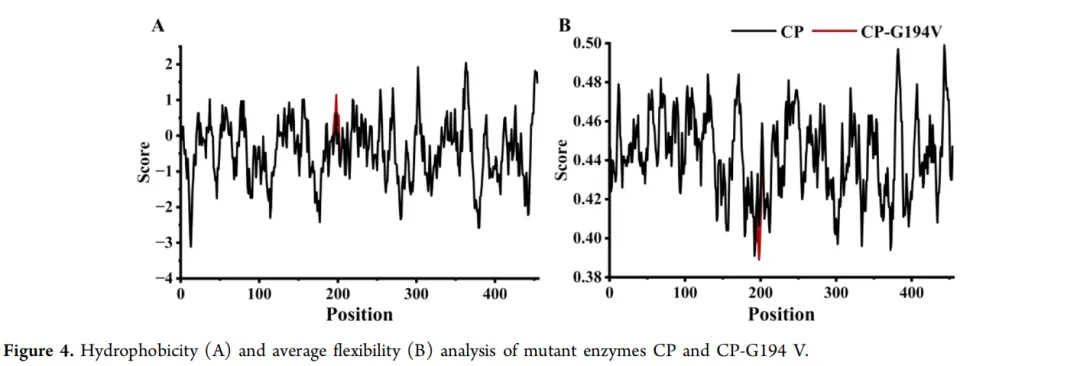
4.1.2. 野生型及其突变体酶的疏水性与柔韧性分析为了深入探究突变酶热稳定性增强的分子机制,利用在线软件ProtScale分析了突变体的疏水性和柔韧性。结果显示,与突变酶CP相比,Gly194残基的突变导致突变酶CP-G194V中Gly194−Gln202区域的疏水性增加(图4A),柔韧性降低(图4B)。观察到热稳定性与疏水性呈正相关,与柔韧性呈负相关。因此,突变酶CP-G194V热稳定性增强的分子机制涉及氨基酸疏水性的升高和柔韧性的降低。
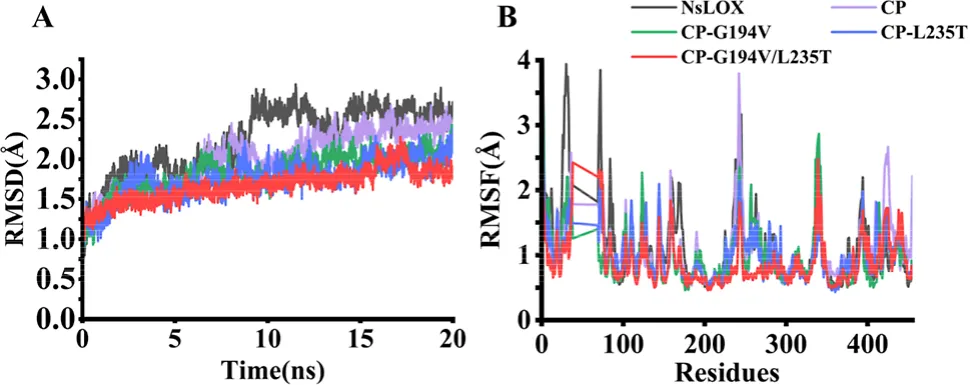
4.1.3. 野生型及其突变体酶的分子动力学模拟分析为了从动力学角度进一步分析突变酶热稳定性增强的分子机制,以野生型NsLOX晶体结构为参考,构建了突变酶的三维结构模型。随后,利用AMBER 18软件对突变酶CP-G194V、CP-L235T和CP-G194V/L235T以及野生型的结构模型进行了分子动力学模拟。突变酶CP-G194V、CP-L235T和CP-G194V/L235T的平均RMSD值分别为1.829、1.757和1.673 Å,均低于野生型(2.239 Å)和突变酶CP(1.997 Å)。这表明突变酶CP-G194V、CP-L235T和CP-G194V/L235T表现出更高的结构刚性和更稳定的构象。值得注意的是,野生型在8 ns处出现一个显著尖峰,表明其在此温度下高度不稳定(图5A)。此外,与野生型相比,突变酶CP-G194V/L235T中特定区域(如Met1−Glu94、Pro102−Asp292、Ile302−Ser335、Thr348−Val359和Asn371−Gly401)的RMSF值显著降低。总体而言,突变酶CP-G194V/L235T在所有氨基酸残基中79.39%的位置上均表现出降低的RMSF值(图5B)。热波动区域分布的减少意味着突变酶具有更高的刚性和稳定性。总之,突变酶CP-G194V、CP-L235T和CP-G194V/L235T热稳定性的增强可归因于分子水平上结构刚性的增加和柔韧性的降低。另外两次独立的模拟得出了一致的趋势,证实了结果的可靠性。
4.2. 突变酶对LA比活力提高的机制解析
4.2.1. 野生型及其突变体酶的空间位置分析
使用AutoDockTools-1.5.7对NsLOX及其突变体与底物LA进行分子对接。对接结果显示,所有结合分数均低于−5 kcal/mol,表明对接构象可靠。此外,野生型与突变体结构的整体折叠高度相似,NsLOX与CP之间的均方根偏差为1.644 Å(389对389个原子),CP与CP-G194V、CP-L235T和CP-G194V/L235T相比的RMSD值分别为1.603 Å(381对381个原子)、1.775 Å(366对366个原子)和1.834 Å(385对385个原子)。这些微小的构象变化与其他酶的观察结果一致。
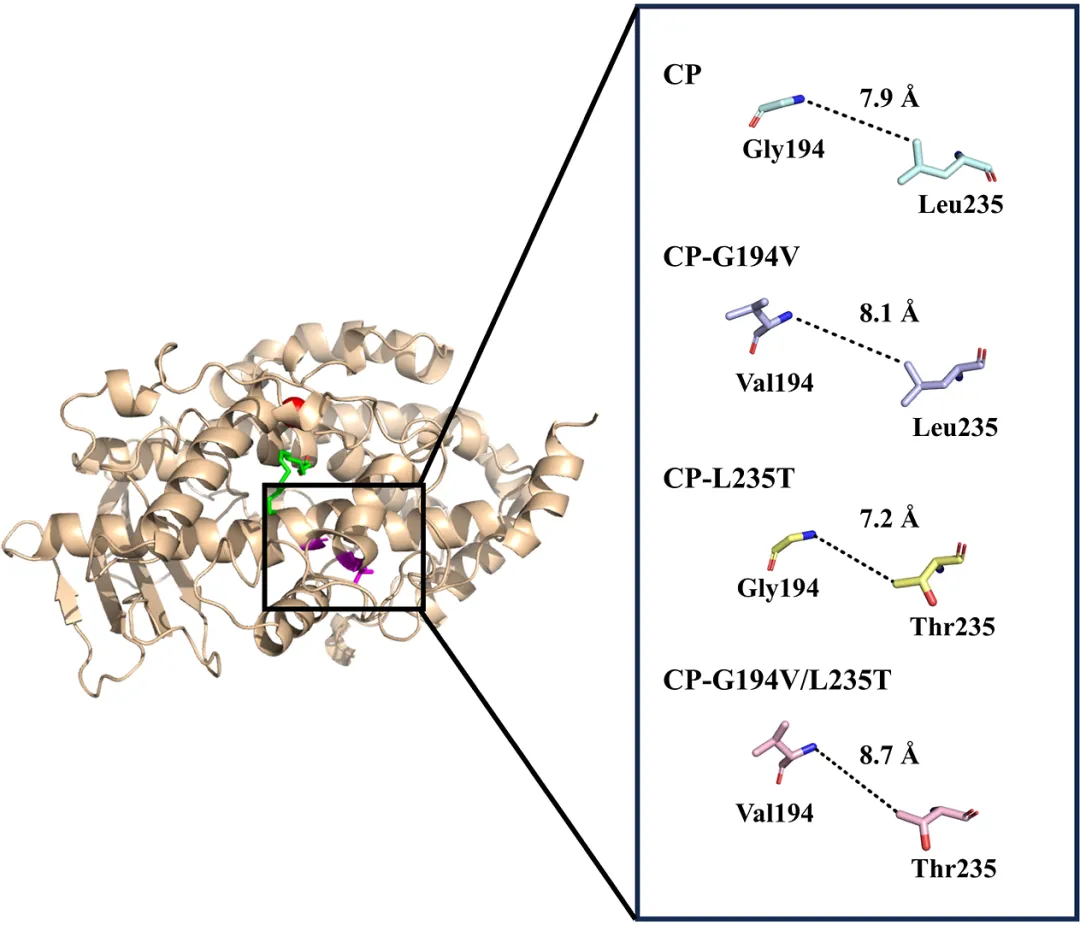
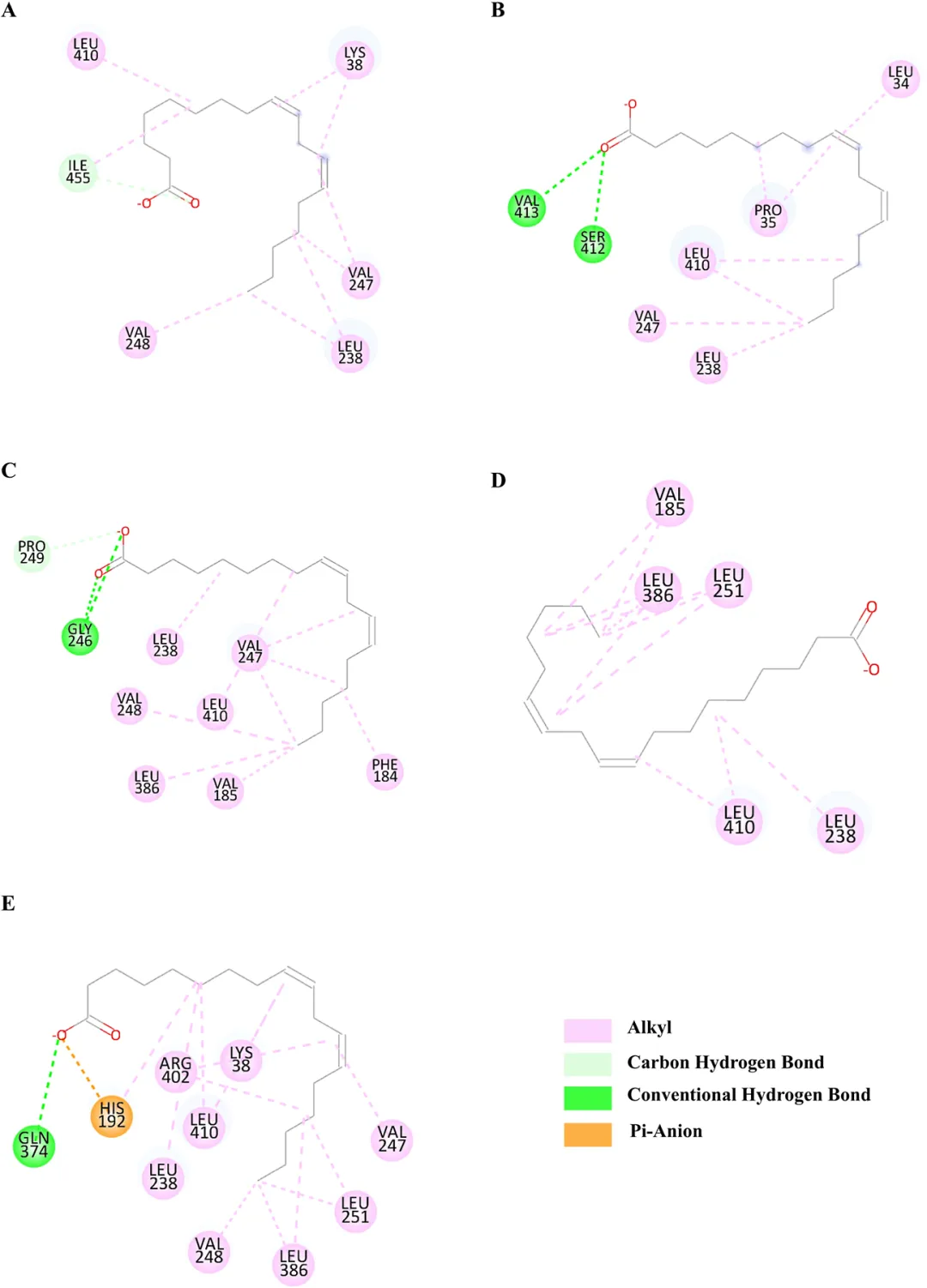
为了探究突变引起催化活性显著变化的原因,分析了突变位点之间的空间距离。Gly194和Leu235位于形成NsLOX催化口袋的两个α-螺旋上(图7)。在突变体CP中,Gly194和Leu235之间的距离为7.9 Å。当Gly194突变为Val(CP-G194V)时,距离增加至8.1 Å。将Leu235突变为Thr(CP-L235T)导致距离为7.2 Å,而双突变体(CP-G194V/L235T)显示距离为8.7 Å。生化数据表明,催化活性按以下顺序递减:CP-G194V/L235T > CP-G194V > CP > CP-L235T。这一趋势与Gly194和Leu235之间的距离相关:距离越大,催化活性越高。这可能归因于突变导致两个α-螺旋发生微妙的位移,从而使结合口袋扩大,有利于底物LA的进入和结合。
4.2.2. 野生型及其突变体酶的底物通道与口袋分析底物进入通道的结构特征显著影响催化活性、底物特异性甚至蛋白质稳定性。因此,对分子通道的深入分析对于理解和调控酶及其他蛋白质的功能特性至关重要。通常,具有更短通道和更宽瓶颈半径的酶展现出更易接近的催化位点。此外,可成药性评分——一个指示口袋结合类药物分子可能性的指标——为进一步了解配体结合潜力提供了见解。为了进一步研究突变引起的酶活性显著变化,使用Caver Web 2.0对NsLOX及其突变体进行了计算分析。通过检查每个变体的瓶颈半径和可成药性评分,我们发现较大的瓶颈半径有利于亚油酸(LA)的进入,表明大体积底物需要更宽敞的通道瓶颈。此外,观察到可成药性评分与酶活性之间呈正相关:可成药性值越高,催化效率越高,这进一步印证了底物结合亲和力的提高增强了催化性能的观点。
4.2.3. 野生型及其突变体酶的活性中心相互作用网络分析为了进一步研究酶活性的变化,分析了突变体与野生型酶之间活性中心相互作用网络的改变。使用Discovery Studio Client分析了酶与底物LA之间的相互作用力。在野生型NsLOX中,LA与Lys38、Val247、Val248、Leu238和Leu410形成烷基相互作用,并与Ile455形成一个氢键(图8A)。在CP突变体中,LA与Leu34、Pro35、Val247、Leu238和Leu410发生烷基相互作用,同时与Val413和Ser412形成强氢键。与野生型相比,虽然失去一个烷基相互作用,但与LA的羧基形成了一个额外的氢键(图8B)。在CP背景下将Gly194突变为Val(CP-G194V)时,LA与Val185、Val247、Val248、Leu238、Leu410、Leu386和Phe184建立烷基相互作用,并与Gly246和Pro249形成强氢键。与CP相比,这增加了两个烷基相互作用(图8C)。在CP-L235T突变体(Leu235突变为Thr)中,LA与Val185、Leu251、Leu386、Leu410和Leu238形成烷基相互作用。相对于CP,烷基相互作用以及与LA羧基的氢键均减少(图8D)。在CP-G194V/L235T突变体中,LA参与与Lys38、Val247、Val248、Leu238、Leu410、Leu251、Leu386和Arg402的烷基相互作用,与Glu374形成一个强氢键,并且值得注意的是,与His192形成一个π-阴离子相互作用(图8E)。与CP、CP-G194V和CP-L235T相比,该突变体表现出更多的烷基相互作用。结合NsLOX及其突变体的催化活性数据,可以观察到,与LA的烷基相互作用数量越多,稳定LA羧基的相互作用越强,催化活性就越高。先前的研究表明,底物进入底物通道的方向是决定LOX催化不饱和脂肪酸氢过氧化反应特异性的因素之一,正确的底物定位对于有效的LA催化至关重要[43]。此外,与CP相比,CPV和CPVT都形成了更疏水的底物通道,这有利于疏水性底物进入催化活性中心。相比之下,CPT既缺乏定位LA的关键残基,也缺乏更疏水的底物通道,导致活性略有降低。
本研究中突变体能够同时实现热稳定性和催化活性增强的内在机制,源于三个层面的协同效应。
研究发现,G194V和L235T突变能够对形成催化口袋的α-螺旋进行精细调控,将关键残基之间的距离从7.9 Å(CP)增加到8.7 Å(CP-G194V/L235T)。这种适度的空间扩张,加上底物通道瓶颈半径的增加,被证明能够协同优化疏水性底物LA的进入和结合,从而解释了比活力提高的趋势。这表明,对活性口袋周围残基的理性设计能够在保持整体构象稳定性的同时,赋予底物识别所需的局部适应性。
突变体在局部形成了更强的疏水相互作用网络(图6),例如Val194与Val190、Leu267等残基之间的疏水接触。这些局部“锚定”效应被认为能够降低构象熵,并传递至整体结构。分子动力学模拟证实,所有耐热突变体的RMSD值均降低(图5A),表明局部疏水相互作用的增强能够有效转化为整体构象刚性的提升。
4.3.3. 相互作用网络的系统优化
催化活性的提高直接归因于底物结合模式的优化:最具活性的突变体CP-G194V/L235T被发现与LA形成最广泛的烷基相互作用,并引入了一个独特的π-阴离子相互作用(图8)。同时,蛋白质的静电性质得到改善,观察到双突变体整体偶极矩显著降低。这反映了更均匀的电荷分布和减少的分子内静电应变,为构象稳定性提供了热力学基础。
因此,本研究揭示了一种机制:通过在活性口袋周围进行精确突变,可以同时实现局部疏水相互作用的协同增强、整体构象刚性的改善、底物结合的优化以及静电平衡的提升。这种多方面的协同效应使酶能够同时克服传统的活性-稳定性权衡,为工业酶的理性设计提供了依据。
为了进一步评估优异突变酶在实际应用中对馒头品质的改善效果,采用质地剖面分析(TPA)方法,系统测定了包括硬度、粘附性、弹性和咀嚼性在内的关键质构特性。结果表明,在相同酶活添加量条件下,与添加野生型酶的样品相比,添加NsLOX突变体CP-G194V/L235T的馒头表现出更低的硬度、更高的弹性、更低的粘附性和更低的咀嚼性。此外,其整体品质优于含有大豆LOX的商业改良剂。在NsLOX处理组与空白对照组及商业大豆LOX制剂组之间,观察到硬度和咀嚼性存在显著差异。NsLOX处理对馒头弹性的影响与空白对照组相似,这一趋势与大豆LOX研究报告的结果一致。此外,NsLOX与商业大豆LOX处理在弹性恢复力方面未检测到显著差异,表明NsLOX突变体CP-G194V/L235T作为面粉改良剂具有优异的综合特性。这进一步证明了热稳定性改善的LOX具有增强的应用潜力。
提高蛋白质热稳定性是酶工程的核心目标之一。具体到LOX,以往的研究方法主要集中于共有序列设计、柔性区域截短及其他分子修饰。例如,Qian等人采用生物信息学指导的共有序列方法获得了热稳定性提高的LOX变体,而Lu等人则通过改造柔性区域或将酶与自组装两亲肽融合来增强LOX的稳定性。这些方法虽然在某些情况下有效,但通常涉及广泛的突变扫描、庞大的实验文库,且对选择目标残基的结构或动力学依据有限,未能系统评估酶的整体结构稳定性景观。除了这些传统方法,常用的提高蛋白质热稳定性的策略还包括二硫键设计、分子对接、量子力学和分子动力学模拟分析。在这些方法中,分子动力学模拟分析脱颖而出,成为一种精确、直接且高效的技术。与静态结构分析不同,分子动力学模拟可以更详细地洞察蛋白质的功能构象。通过利用MD模拟,研究人员可以精确定位蛋白质中的动态柔性区域。这些区域的变化,特别是在高温条件下,能显著影响蛋白质的稳定性和活性。当进行高温模拟时,由于温度升高,蛋白质内的分子运动加速,导致不稳定区域去折叠,蛋白质结构变得松散[51]。因此,比较高温模拟前后的结构差异,可以快速识别出柔性区域[14,15]。然而,通过这种方法识别出的柔性区域通常包含大量的氨基酸残基,这大大增加了后续突变筛选的工作量。
越来越多的研究表明,构建更小、更聚焦的突变文库可以极大提高实验效率,而精心设计的文库能显著增加获得有益突变的可能性。Reetz强调,许多研究小组正在开发生成小型、智能突变文库的方法,以克服分子修饰中固有的劳动密集型步骤。类似地,Zhou等人证明,理性设计可以提高获得有益突变的可能性,同时节省大量时间和精力;Qin等人[54]进一步强调,减少热点数量和突变文库规模是实现高效酶设计的关键原则。因此,有必要整合多种互补策略进行理性酶设计,以提高筛选效率。
此外,进行多序列比较是提高酶热稳定性的另一种有效方法。通过鉴定在热稳定性中起关键作用的氨基酸位点,并通过序列比较将其替换为耐热酶中的相应残基,可以取得显著改善。在本研究中,通过分子动力学模拟、多序列比较和保守性分析鉴定了关键氨基酸残基。通过定点饱和突变和组合诱变的结合,成功开发了突变体CP-G194V/L235T,其热稳定性显著增强。该酶在50°C的半衰期为389.08 ± 36.62分钟,比CP突变体长124.72分钟,是野生型的45.29倍。此外,其Tm值比野生型高8.04°C。这些结果表明,采用多组合策略是提高酶热稳定性的有效途径。与传统方法相比,该策略极大地缩小了需要筛选的突变文库规模——从八个区域的134个氨基酸残基减少到仅五个合理识别的热点残基——从而大大提高了成功率和整体效率。这些结果证明了我们应用的结构导向组合策略在解决该LOX的权衡问题上的有效性。
酶的催化活性和热稳定性是评估其工业应用性的两个关键性能指标。具有高催化活性和良好热稳定性的酶在工业环境中更具竞争力,拥有更大的应用潜力。研究表明,酶的催化效率主要取决于其活性位点的结构柔性,而酶分子的整体稳定性则与其三维结构的刚性密切相关。这种对结构和功能的双重需求导致了酶工程中普遍存在的“稳定性-活性”权衡效应。这一科学挑战已成为限制工业酶性能提升的关键瓶颈。突变酶CP-G194V的比活力显著高于野生型和突变酶CP,达到215881.25 ± 5024.47 U/mg。另一方面,突变酶CP-L235T的比活力(65472.18 ± 5525.32 U/mg)略低于野生型。然而,该突变并未对组合突变酶CP-G194V/L235T的比活力产生不利影响。有趣的是,CP-G194V/L235T中的突变组合并未降低其比活力,反而协同地将酶的比活力提高至225613.6 ± 11944.25 U/mg,是野生型的2.89倍。总之,采用多突变策略可以有效提高酶的热稳定性和催化活性。该方法被证明是克服酶催化活性与热稳定性之间常见权衡的有效途径。
本理性设计策略基于将突变靶向保守性较低的区域以保留催化功能。虽然有效,但这种方法固有限制了研究保守残基潜在稳定性贡献的能力。这些通常被认为对催化不可改变的残基,可能是在进化中选择出来在结构完整性中发挥双重作用的,这为“第二代”工程提供了一个引人注目的途径。未来的工作可以采用祖先序列重建来推断这些残基增强稳定性的历史状态。或者,对活性位点微环境进行靶向深度突变扫描,结合高保真度活性测定,可以系统地检验这一假设,并揭示稳定性和活性可以协同优化的“隐藏”热点,从而超越现代序列保守性的限制。此外,应注意本研究中揭示的G194V和L235T之间的强协同效应是在预稳定的CP(T2C/S36P)骨架中观察到的。虽然这证实了筛选策略在识别高阶稳定性热点方面的有效性,但这种协同效应是否适用于野生型或其他LOX同源物,仍有待未来的实验验证。本研究为此类验证提供了明确的靶点和理论基础。
此外,本研究的一个关键发现是,以亚油酸为底物观察到的显著协同效应模式,在使用ALA时发生了特异性转变。一方面,CP和CP-G194V突变体对ALA的活性也显著增加,表明这些关键残基的替换赋予了一定程度的底物普适性以增强催化效率。另一方面,突变组合的最佳协同模式显示出严格的底物依赖性:在LA体系中表现最佳的CP-G194V/L235T,在ALA体系中的活性显著低于CP-G194V。这一现象深刻揭示,本研究中获得的突变并非简单的“全局激活剂”。相反,它们很可能通过重塑活性位点的局部微环境,对不同底物的催化过程产生差异性影响。LA(18:2,n-6)和ALA(18:3,n-3)碳链中双键数量和位置的差异,可能导致它们在与活性位点结合时采取不同的构象和电子分布。因此,同一组突变对不同底物可产生显著不同的协同效应。上述发现不仅阐明了酶-底物相互作用的精确性和特异性,也指出了一个更具挑战性和前景的研究方向:能否通过理性设计改造活性口袋的兼容性,开发出对多种多不饱和脂肪酸底物均能保持高效协同催化性能的下一代工业酶?这将是实现复杂天然油脂底物高效、多功能生物转化的关键。
脂氧合酶的工业应用要求酶具有高产量、高纯度、酸碱耐受性、热稳定性和催化活性。在此背景下,突变体CP-G194V/L235T不仅表现出增强的热稳定性,而且在5−25°C的温度范围内保留了超过70%的相对活性。相比之下,野生型NsLOX在相同的低温条件下仅保持约40%的相对活性。这一性能与实际的面粉加工过程特别相关。例如,在添加阶段——LOX通常在此阶段被加入到面粉中——工业食品设施的环境温度通常保持在16至28°C之间。在此温和范围内操作有助于保持产品的营养和感官品质,同时降低能耗和环境影响。因此,我们突变体的高温活性谱与实际工业条件非常吻合。将我们的工程酶与商业替代品进行比较也很重要。大多数商业LOX制剂来源于大豆粉或粗大豆提取物,它们存在若干局限性:LOX比含量低、存在多种具有不同催化行为的同工酶(LOX-1、LOX-2和LOX-3)、基质组成复杂,以及可能影响最终产品品质的深色。在大豆LOX同工酶中,只有LOX-1在低温下表现出显著的活性。例如,当使用大豆LOX在2°C、以亚麻酸为底物的10 L反应器中时,13-氢过氧化物的生产率仅达到60 g/L·h。此外,大豆LOX的比活力为0.98 μM/min·μg,最适反应温度为35°C,催化效率(kcat/Km)为0.48 s⁻¹·μM⁻¹。NsLOX突变体CP-G194V/L235T在工业相关条件下表现出优异的化学耐受性,有效抵抗了工业催化中常见的化学胁迫。通过馒头TPA进行的进一步应用评估显示,在酶活添加量相等的情况下,含有突变体CP-G194V/L235T的样品在硬度、粘附性和咀嚼性方面比用野生型酶制备的样品表现更佳,同时其整体品质也超过了基于商业大豆LOX的改良剂。这些特性凸显了其作为传统大豆LOX在面粉增白和面团调理应用中的优质替代品的潜力。
已有研究表明,通过降低表面和活性空腔残基的柔韧性可以增强酶的热稳定性。这表明降低柔韧性是提高酶热稳定性的关键因素。突变酶CP-G194V/L235T的平均RMSD值为1.673 Å,低于野生型(2.239 Å)和突变酶CP(1.997 Å)。此外,与野生型相比,突变酶CP-G194V/L235T中多个区域(包括Met1−Glu94、Pro102−Asp292、Ile302−Ser335、Thr348−Val359和Asn371−Gly401)的RMSF值显著降低。值得注意的是,突变酶CP-G194V/L235T中79.39%的氨基酸残基的RMSF值降低。较低的RMSF值表明分子结构更稳定,原子距离波动更小。多种结构因素影响蛋白质的热稳定性,如静电相互作用、盐桥、二硫键、氢键、疏水相互作用、二级结构和氨基酸组成。氢键和疏水相互作用尤其关键,前者有助于蛋白质稳定性,后者为蛋白质折叠提供能量。对突变酶CP-G194V的疏水性和柔韧性分析表明,Gly194残基的突变导致Gly194−Gln202区域内疏水性增加、柔韧性降低。含有更高比例疏水残基(如Val、Ile和Pro)的嗜热蛋白可增强蛋白质的疏水性、刚性,从而提高热稳定性。对突变酶疏水相互作用的研究表明,CP-G194V和CP-L235T分别与周围氨基酸形成了四个新的疏水相互作用。这些疏水相互作用是维持蛋白质折叠的主要驱动力,对蛋白质稳定性的贡献约为64%。疏水相互作用的增加增强了酶分子的构象熵,导致突变酶的三级结构更稳定。这一机制在提高突变酶的整体热稳定性方面起着至关重要的作用。此外,蛋白质表面的电荷特性与其改变形状的能力密切相关。已知带负电的表面会显著增强蛋白质的柔韧性,而带正电和电中性的表面则倾向于增加刚性。对突变酶静电势的检测显示,残基Gly194和Leu235的突变分别导致蛋白质表面变得更中性和更正,从而增加了刚性。这种增强有助于突变酶CP-G194V和CP-L235T保持高度稳定的构象。研究结果表明,蛋白质表面具有正电或电中性势的残基比具有负电势的残基表现出更大的构象刚性。研究表明,底物进入底物通道的方向是决定LOX催化不饱和脂肪酸氢过氧化反应区域/立体特异性的因素之一,而底物的正确定位对于确保亚油酸的有效催化至关重要[43]。此外,突变诱导了活性中心附近两个α-螺旋的轻微位移,从而扩大了底物通道,增加了底物可及性。尽管这两个突变位点不直接参与底物结合,但它们引起了活性中心相互作用网络的扰动。
在野生型和CP突变体中,Leu235是一个疏水残基,其侧链与周围的疏水残基(如G194区域内的残基)形成稳定的疏水相互作用网络。该网络对于维持活性口袋正确的局部结构和疏水特性至关重要。当突变为体积更大且含有极性羟基的苏氨酸(Thr)时,Thr235的侧链很可能引入轻微的空间冲突,并迫使局部环境适应一个亲水基团。这破坏了原本紧凑、疏水的局部环境。如图8D所示,与其他突变体相比,CP-L235T与底物LA的相互作用(包括烷基相互作用和氢键)更少,直接反映了其结合口袋在容纳和稳定疏水性LA底物方面的效率降低。此外,空间位置分析(图7)显示,L235T突变将两个α-螺旋之间的距离缩短至7.2 Å(CP中为7.9 Å)。这表明该突变可能“过度收紧”或限制了构成活性口袋的α-螺旋的构象柔性。具有一定柔性的活性口袋通常是底物进入、诱导契合和产物释放所必需的。CP-L235T可能使该区域过于刚性或将其锁定在不利于最佳底物结合的构象中,从而降低了催化效率。这一发现与该突变体相对较窄的底物通道瓶颈半径是一致的。
准确理解CP-L235T的负面影响,恰恰揭示了双突变体CP-G194V/L235T中协同效应的精妙之处。提出,G194V突变有效地“挽救”并“利用”了L235T突变诱导的变化:G194V突变引入了一个更大的疏水侧链(Val),这本身倾向于向外推挤α-螺旋,扩大底物口袋(如CP-G194V中距离增加到8.1 Å所示)。在CP-G194V/L235T双突变体中,G194V的这种“扩张”效应正好补偿了L235T可能引起的构象收缩和疏水环境破坏。它不仅抵消了不利的空间效应,还与Thr235协同作用,共同将活性口袋重塑为一个具有更大尺寸(距离增至8.7 Å)、更优疏水性(更多烷基相互作用;图8E)以及能够形成新的关键相互作用(如与His192的π-阴离子相互作用)的理想构象。简而言之,L235T单独存在时是有害的,因为它破坏了局部结构和动态平衡;然而,当与G194V配对时,后者提供了一种结构“补偿力”,将整个系统推向一个优于任一单突变体的功能状态。这就解释了为什么这两个突变一起产生了“1 + 1 > 2”的协同增强效应。
必须明确指出,目前提出的“补偿力”机制主要源于结构动态变化的观察和相关性分析,本质上构成了一个描述性的机制模型。尽管突变位点间距离的协同变化为该模型提供了有力支持,但尚未进行定量能量计算(如结合自由能或相互作用能分析)来从热力学上直接论证Val194和Thr235侧链间的相互作用如何稳定目标构象。这一局限性限制了该模型在未来指导精确理性设计方面的预测能力。本研究的主要价值在于首次实验发现了G194V和L235T之间的协同增强效应,并基于现有结构生物学工具构建了一个可检验的机制假说,为后续探索奠定了基础。为了推动该模型从“现象描述”走向“精确预测”,必须引入更深入的计算能量验证。未来的工作可集中在:(1) 采用MM/PBSA或自由能微扰方法,定量评估每个突变对不同构象状态之间自由能差的贡献;(2) 利用QM/MM计算,在电子结构水平上解析突变簇如何精细调控活性中心的局部相互作用网络;(3) 在更长的分子动力学模拟基础上,进行更详细的动态分析,如构象熵和残基相互作用网络分析。通过这些计算方法,有望在能量水平上直接验证或修正“补偿力”假说,从而建立突变与功能之间的定量关系。最终,这将推动脂氧合酶及其他工业酶的理性设计迈向可预测性增强的新阶段。
总之,本研究利用多重组合策略成功开发了突变酶CP-G194V/L235T,其催化活性和热稳定性显著增强。CP-G194V/L235T在50°C下的半衰期为389.08 ± 36.62分钟,Tm值为67.85 ± 0.40°C,比活力为215,881.25 ± 5024.47 U/mg。这是首次通过综合运用分子动力学模拟、多序列比较、结构与保守性分析以鉴定关键氨基酸残基,并结合定点饱和突变与组合诱变,实现增强NsLOX热稳定性和催化活性的报道。进一步的二级结构、序列、分子动力学模拟和三维结构分析表明,突变体CP-G194V/L235T表现出增加的氨基酸疏水性、降低的柔韧性、增强的结构刚性、强化的疏水相互作用以及改善的表面正电和电中性势,从而提高了其热稳定性。此外,增强的酶-LA相互作用,特别是锚定LA羧基的相互作用,导致了优化的底物结合和定位。这些针对性的调整维持了酶的结构和催化完整性,从而在不损害催化活性(在某些情况下甚至增强)的前提下,实现了热稳定性的显著提高。本研究为解决酶催化活性与热稳定性之间的权衡问题提供了一种精确有效的策略,为提高食品工业应用中酶的热稳定性提供了理性高效的方法和宝贵的见解。