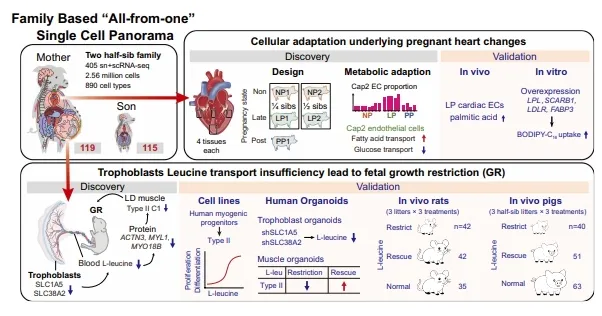
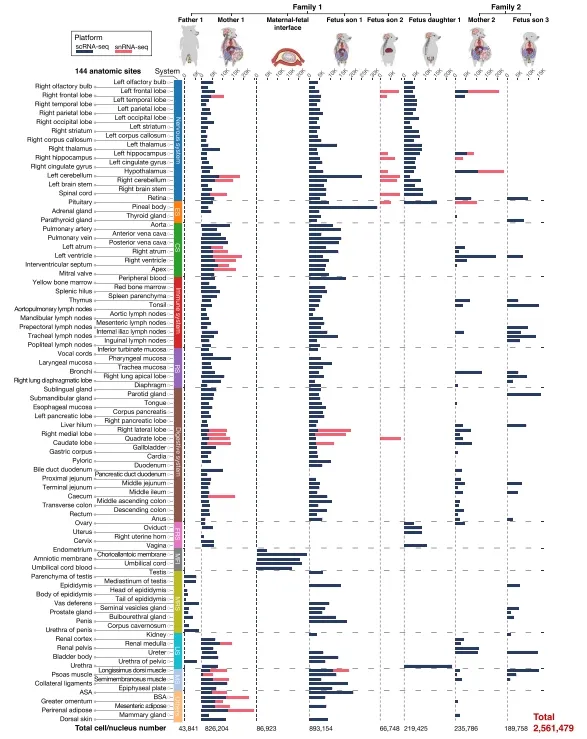
研究团队从一头妊娠110天的母猪(F₁M₁)和她的一个公胎(F₁S₁)身上,分别采集了119个和115个组织,覆盖神经、循环、免疫、生殖、消化等12大系统。所有组织在16小时内完成解离和测序,最终获得2,561,479个高质量单细胞。【图1】
图1.两个猪家系 “单一来源” 单细胞全景图谱概述
复现建议:取组织前先固定好解离方案,不同器官酶配比差异大。我们提前配好足量消化液,120人分头行动,16小时内完成275个样本。建议先做预实验摸条件,不然样品容易降解。
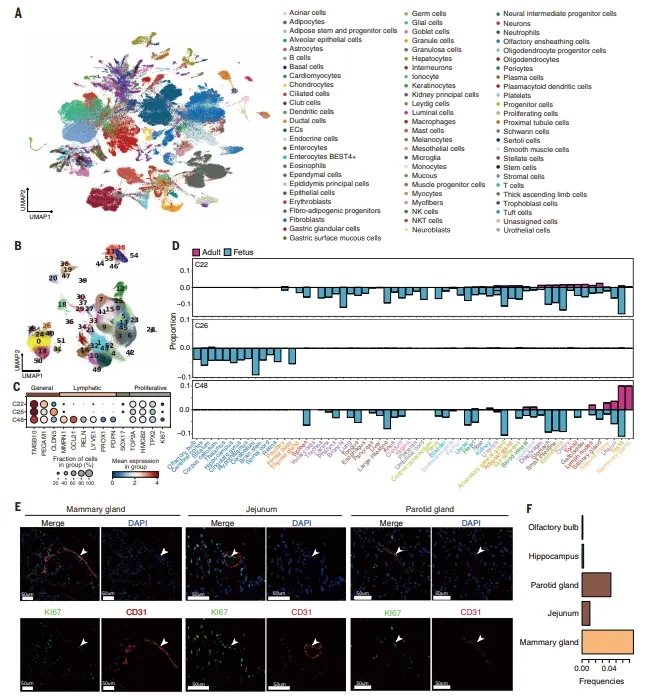
全局UMAP展示76种主细胞类型,内皮细胞进一步分为55个亚群。增殖内皮C22和C26高表达TOP2A等标志,C26仅见于胎儿大脑。免疫荧光证实乳腺、空肠等成人组织中存在增殖内皮。【图2】
图2.构建猪胎儿与成年个体全身器官的一体化单细胞图谱,并解析全身器官内皮细胞的分布特征
复现建议:鉴定增殖内皮时,CD31和KI67共染背景高,建议用TSA放大信号。内皮亚群定义需结合动脉/静脉/淋巴标志基因,我们参考了小鼠内皮图谱,跨物种对比能提高可信度。
59个器官的内皮亚型UMAP共鉴定412个内皮亚型(动脉、毛细血管、静脉、淋巴等)。183个免疫基因在超80%亚型中普遍表达,富集于抗原呈递和白细胞迁移。脑内皮几乎不表达CD36和MHC II基因。【图3】
图3.59 种器官中内皮细胞的器官内异质性与免疫调节功能多样性
复现建议:每个器官内皮单独聚类再整合,避免大整合掩盖低频亚型。免疫基因集用KEGG pathway,清道夫受体从HGNC批量获取。跨器官比较时注意批次校正,我们用了Harmony。
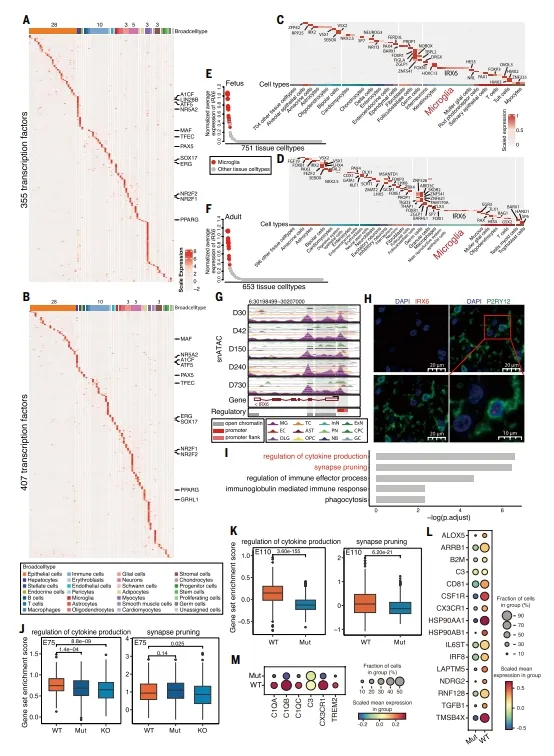
利用“一人一图”的优势,研究识别出355个胎儿和407个成年细胞类型特异性转录因子。其中,IRX6在所有11个脑区的小胶质细胞中高表达,而在其他细胞中几乎不表达。通过基因编辑猪和snATAC-seq验证,IRX6调控突触修剪和细胞因子产生通路。这是首次在大型哺乳动物中确认IRX6为小胶质细胞的核心转录因子。【图4】
复现建议:细胞特异TF用tau>0.9筛选。验证IRX6功能需构建基因编辑猪,我们做了起始密码子突变和exon4敲除两种。snATAC-seq用ArchR分析,注意不同细胞类型峰背景噪声差异大。
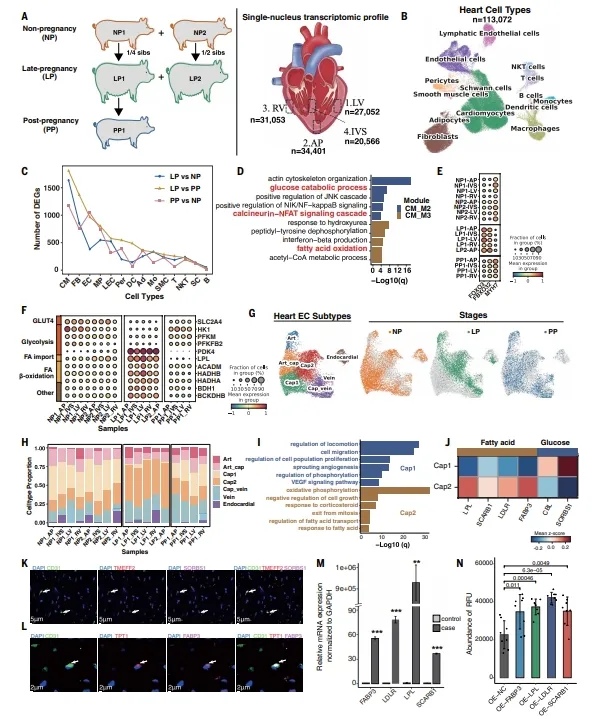
妊娠心脏snRNA-seq发现Cap2毛细血管亚群在孕期富集,产后消失。Cap2高表达脂肪酸转运基因,Cap1高表达葡萄糖转运基因。FOXO3和FBXO32孕期上调,抑制钙调磷酸酶-NFAT通路,避免病理性肥厚。心肌细胞代谢基因PDK4、ACADM等孕期上调,糖酵解基因下调。【图5】
图5.猪心脏在 NP、LP 和 PP 阶段的动态细胞及功能适应性变化
复现建议:妊娠心脏样本难获取,建议用半同胞母猪对照减少遗传干扰。Cap2和Cap1的分选可用FABP3和SORBS1抗体。脂肪酸摄取用BODIPY-C16,葡萄糖功能通过敲低转运基因后测代谢物验证。
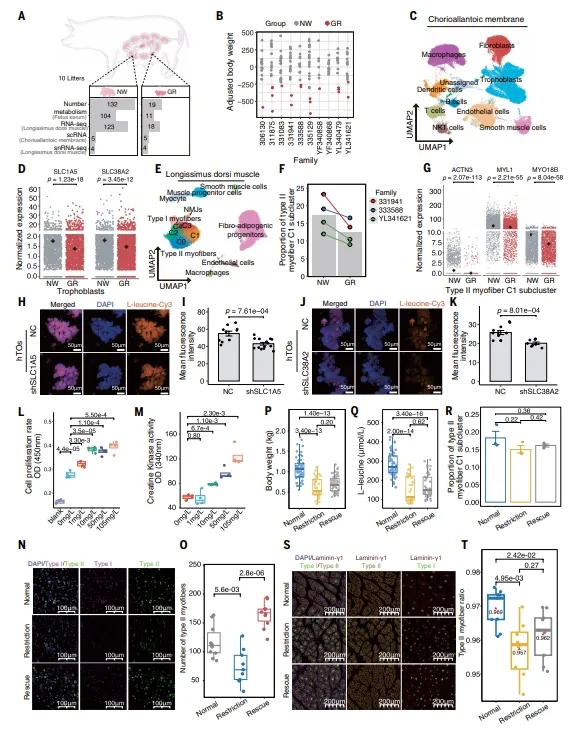
生长受限胎儿胎盘滋养层中SLC1A5和SLC38A2表达降低,胎血亮氨酸减少。肌肉IIC1亚簇比例下降,ACTN3等基因表达降低。人滋养层类器官、肌原祖细胞及猪/鼠体内补充实验验证亮氨酸缺乏限制肌纤维发育。【图6】
复现建议:FGR模型利用同窝内自然差异即可,不需额外造模。滋养层类器官用基质胶培养,亮氨酸摄取用Cy3标记的L-亮氨酸。肌肉类器官构建时注意基质胶配方和分化时机,挽救试验时间点很关键。
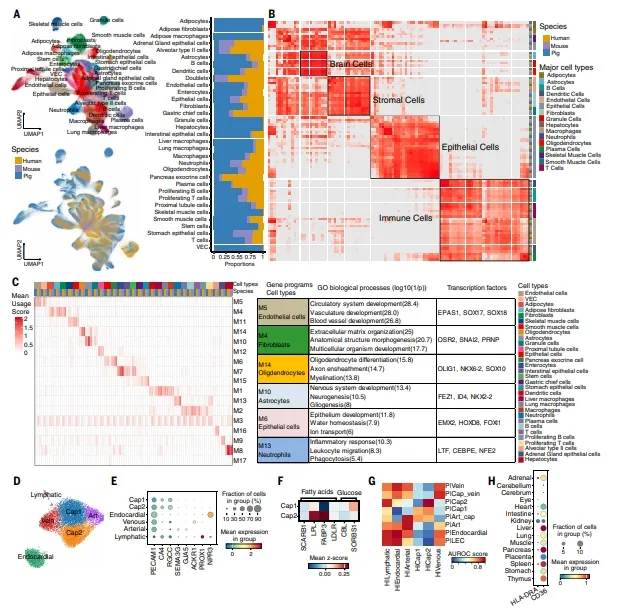
整合人、小鼠、猪单细胞数据,免疫、内皮、基质、脑细胞跨物种高度保守。人心肌也存在Cap1/Cap2亚型,且Cap2高表达LPL/FABP3。人胎儿脑中HLA-DRA和CD36表达极低,与外周组织一致。【图7】
图7.对比分析结果表明,猪、人类和小鼠的细胞类型以及基因表达模式在物种间具有保守性
复现建议:跨物种整合前,先把基因名统一为同源基因。cNMF分析建议设多次随机种子,选稳定程序。比较人心肌内皮时,直接用公开数据集,复现我们的分析流程即可。