国家重点研发计划|中国农业大学闫巧娟教授:大曲中拜赖青霉发酵产单宁酶条件优化、纯化及酶学性质分析
- 2026-06-03 23:28:21


EI收录,
入选中国科技期刊卓越行动计划

本文获国家重点研发计划项目 2021YFC2103004。
摘要
单宁广泛分布于植物的叶、皮、根及果实等组织中,其分子中含有大量的酚羟基,能够与金属离子和蛋白质形成稳定的络合物。单宁酶(EC 3.1.1.20)可以催化单宁中酯键的水解,生成没食子酸和其它酚类物质,在植物的代谢调控中具有重要作用,同时在工业领域中也具有广泛应用,例如用于高效制备没食子酸等。单宁酶在动物、植物和微生物中普遍存在,其中微生物是单宁酶最主要的来源,如细菌、酵母菌和丝状真菌(如曲霉属和青霉属)均存在单宁酶。野生型微生物发酵产单宁酶的生产水平通常较低,如烟曲霉(Aspergillus fumigatus)AN1和皮壳青霉(Penicillium crustosum)AN3的产酶水平分别为4.3 U/mL和8.4 U/mL。优化野生型微生物的发酵条件可显著提高产单宁酶水平,例如通过优化黑曲霉(Aspergillus niger)J-T18和灰绿青霉(Aspergillus glaucus)的发酵产单宁酶条件,产单宁酶水平得到明显提升。多数单宁酶在高温和酸性环境下的稳定性差,难以满足不同的应用需求,如地丝菌属酵母(Geotrichum cucujoidarum)来源单宁酶在40 ℃下处理30 min,残留酶活力低于40%;黑绿青霉(Penicillium atramentosum)EZ-ZH190来源单宁酶在pH3.5下处理30 min,残留酶活力小于35%。因此,发掘产酶水平高、酶学性质好的野生型微生物是目前研究单宁酶的重点之一。
大曲是中国白酒酿造过程中不可或缺的传统发酵剂,其核心功能源于复杂的微生物群落及酶系协同作用。微生物通过分泌酶类将原料中的淀粉、蛋白质、脂肪和单宁等大分子物质降解为氨基酸、脂肪酸及酚类化合物等风味前体物质。酿酒原料高粱中丰富的单宁底物可在单宁酶的酶促作用下转化为酚类化合物,如愈创木酚(水果香)、4-乙烯基愈创木酚(花香)和4-甲基愈创木酚(烟熏风味),从而赋予白酒独特的风味特征。已有研究报道,从大曲中分离出产单宁酶菌株烟曲霉HBHF5。大曲中筛选微生物,利用其丰富的微生物资源,为产单宁酶菌株的挖掘开辟了新的方向。
没食子酸是一种天然酚类化合物,存在于多种水果和药用植物中,具有抗氧化、抗炎和抗肿瘤等特性,在食品和制药工业中具有广泛的应用。相比传统化学方法,生物酶法制备没食子酸因其反应条件温和、产物纯度高和产率高以及对环境友好等优势,普遍认为是替代传统化学方法的绿色工艺,成为研究热点。不同底物浓度对酶法制备没食子酸的生成量和产率影响显著,黑曲霉B1401来源的单宁酶水解单宁酸生成的没食子酸浓度随底物量增加先上升后下降,没食子酸产率则呈现下降趋势。汉森隐球菌(Debaryomyces hansenii)来源单宁酶以10和20 g/L单宁酸为底物的没食子酸产率分别为97%和92%,而30 g/L单宁酸为底物的没食子酸产率则降至81%。因此,研究底物浓度对没食子酸生成量和产率的影响,有助于优化酶法制备工艺,提升产物生成量。
本研究旨在从大曲样品中筛选得到一株高产单宁酶的微生物,优化其发酵产酶条件,并评估该酶的酶学性质、水解特性及其在酶法制备没食子酸中的应用潜力。研究充分挖掘了大曲微生物资源,拓宽了产单宁酶的微生物来源,并提供了一种酶学性质优良并能够高效水解单宁酸生成没食子酸的新型单宁酶资源。

结果与分析

2.1 大曲中高产单宁酶拜赖青霉的筛选和鉴定
采用透明圈平板筛选法初筛和甲醇罗丹宁比色法复筛,总计筛选出5株产单宁酶的菌株。其中一株产生较大透明圈的真菌CAU46-1,甲醇罗丹宁比色法复筛的发酵液酶活力为6.2 U/mL,酶活力最高,选定该菌株进行后续研究。
该真菌在PDA培养基上初期为白色绒状菌丝,后期产孢区渐变为青绿色,边缘保留白色菌丝带(图1A)。显微镜(200×)显示该菌菌丝体细长、透明、有隔,分生孢子梗直立,顶端形成典型帚状枝,具单轮生分枝,分生孢子呈球形至亚球形,表面光滑,呈链状排列(图1B)。对照真菌鉴定手册,初步确定该菌株为青霉属。
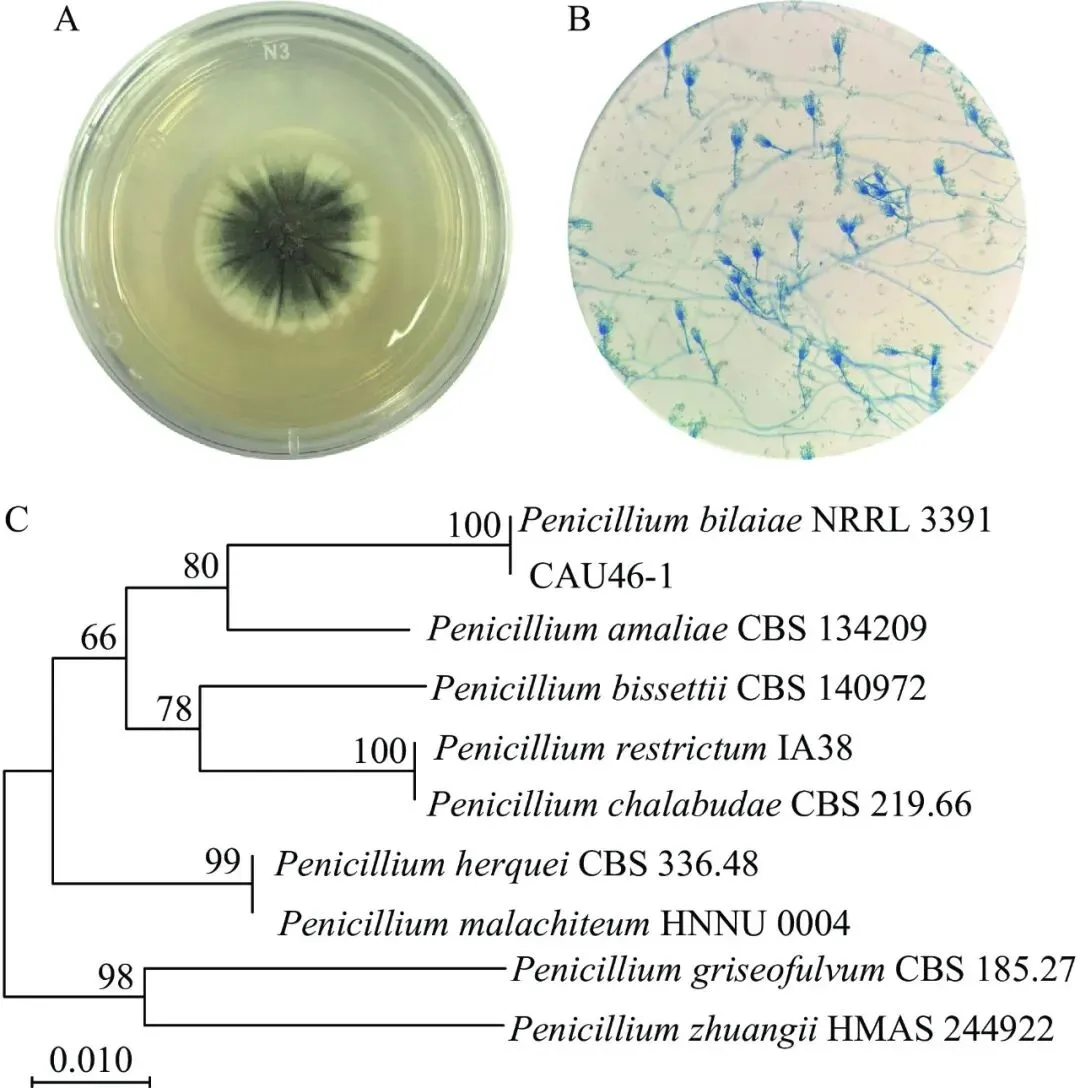
图 1菌株CAU46-1菌落形态(A)、显微特征(B)和ITS序列系统发育树(C)
Figure 1.Colony morphology (A), microscopic characteristics (B) and phylogenetic tree of ITS sequences (C) for strain CAU46-1
对菌株CAU46-1进行ITS基因序列扩增,得到长度为560 bp的ITS区域序列,系统发育树分析表明,该菌ITS序列与拜赖青霉NRRL 3391聚于同一进化支,相似度为100%,与其他青霉物种明显分化,相似度均不高于80%(图1C)。综合菌落形态、显微特征和ITS序列,鉴定该菌为拜赖青霉,命名为拜赖青霉CAU46-1。
2.2 拜赖青霉发酵产单宁酶条件优化
通过单因素实验对拜赖青霉CAU46-1液体发酵产单宁酶的条件进行优化。培养基组分优化结果表明(图2),以单宁酸为诱导物时,拜赖青霉CAU46-1的产酶水平最高,为6.2 U/mL,显著高于没食子酸丙酯和石榴皮粉(P<0.05),无诱导物或以没食子酸为诱导物时该菌不产单宁酶(图2A)。诱导物浓度优化结果表明,发酵液中单宁酸浓度为3%时,菌株产酶水平显著高于其他浓度水平(P<0.05),为10.2 U/mL(图2B)。发酵碳源优化结果表明,以小麦麸皮为碳源时,菌株产酶水平最高,为12.4 U/mL,显著高于其他四种碳源(P<0.05)(图2C)。小麦麸皮浓度为4%时菌株产酶水平最高,达16.2 U/mL,显著高于其他浓度水平(P<0.05),继续提高小麦麸皮浓度,酶活水平显著下降(P<0.05)(图2D)。发酵氮源优化结果表明,无机氮源比有机氮源更利于菌株产酶,其中以氯化铵为氮源时菌株产酶水平最高,为16.9 U/mL(图2E)。氯化铵浓度为0.2%时菌株产酶水平最高(17.0 U/mL),继续提高氯化铵浓度,产酶水平显著下降(P<0.05)(图2F)。发酵参数优化结果如图3所示,该菌最适发酵温度为30 ℃,在此温度下培养3 d,菌株产酶水平为17.3 U/mL,显著高于其他温度水平(P<0.05)(图3A)。不同pH对拜赖青霉CAU46-1液体发酵产单宁酶的影响如图3B,该菌最适发酵pH为5.5,在此pH下培养3 d,菌株产酶水平为20.8 U/mL,显著高于其他pH水平(P<0.05)。在上述优化条件下,拜赖青霉CAU46-1液体发酵产单宁酶历程如图3C,发酵1~4 d时,产酶水平持续提高,发酵4 d后,产酶水平达到最高值(28.7 U/mL)。综上,拜赖青霉CAU46-1液体发酵产单宁酶的最适条件为以3%单宁酸为诱导物、4%小麦麸皮为碳源、0.2% 氯化铵为氮源,在30 ℃、pH5.5下发酵4 d,产酶水平达28.7 U/mL,较优化前提高了3.3倍。
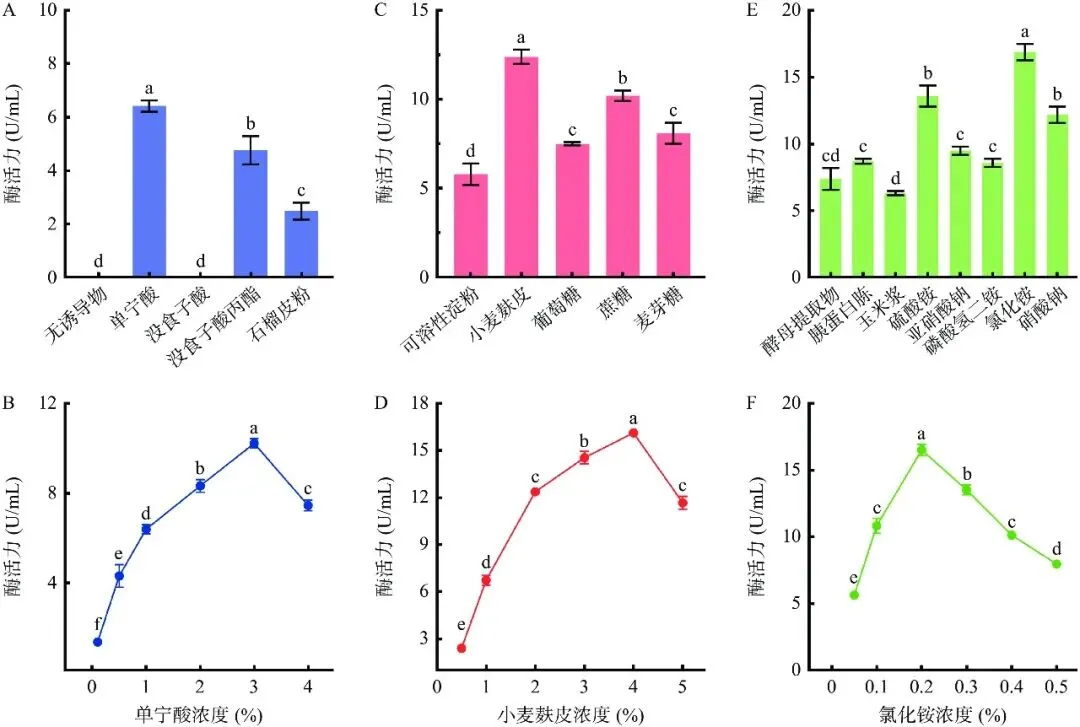
图 2培养基组分对拜赖青霉CAU46-1发酵产酶的影响
Figure 2.Effect of culture medium components on enzyme production of Penicillium bilaiae CAU46-1
注: A:诱导物筛选;B:诱导物浓度优化;C:碳源筛选;D:碳源浓度优化;E:氮源筛选;F:氮源浓度优化;不同小写字母表示有显著性差异,P<0.05,图3同。
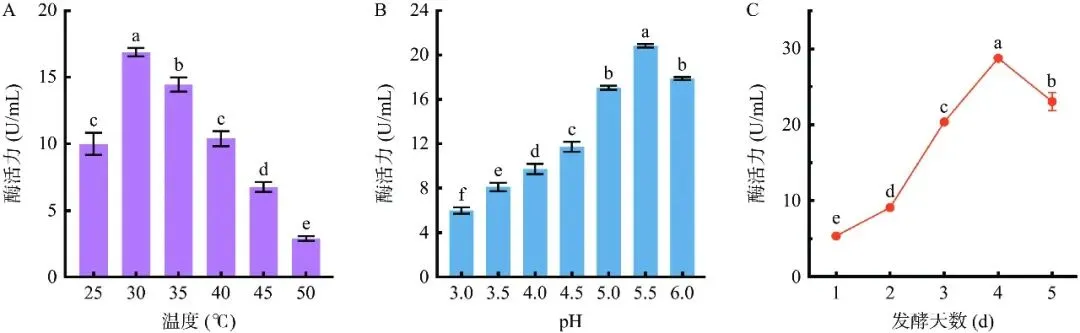
图 3发酵参数对拜赖青霉CAU46-1发酵产酶的影响
Figure 3.Effect of fermentation parameters on enzyme production of Penicillium bilaiae CAU46-1
注:A:发酵温度优化;B:发酵pH优化;C:发酵时间优化。
单宁酶是一种诱导酶,野生型微生物发酵时需要添加单宁或含有单宁的其他物质作为诱导物诱导菌株产酶,多数野生型微生物的最适诱导物为单宁酸。培养基中高浓度单宁酸会导致野生型微生物细胞表面蛋白质变性,损害野生型微生物的新陈代谢和生长,因此多数野生型微生物发酵产单宁酶时单宁酸的浓度在0.5%~3.5%范围内,如黑曲霉AUMC 5614和黄曲霉(Aspergillus flavus)MTCC 3783发酵产单宁酶时,分别以2%和3.2%单宁酸为最适诱导物。小麦麸皮不仅可以作为碳源提供能量,还含有一些单宁和多酚类化合物,促进单宁酶的合成和分泌。除小麦麸皮外,其他富含单宁的农业副产物也常作为野生菌发酵的碳源,如虹色曲霉(Aspergillus versicolor)、木贼镰刀菌(Fusarium equiseti)和桔青霉(Penicillium citrinum)能够利用菠萝皮和金合欢果实作为碳源发酵产单宁酶。氮源是影响野生型微生物产单宁酶的另一关键因素。无机氮源更适合真菌发酵产单宁酶,如黑曲霉J-T18和黑曲霉AUMC 5614分别以硫酸铵和硝酸钠为氮源达到最高产酶水平。产单宁酶野生型微生物的最适发酵温度与菌株的生长环境密切相关,大多在25~40 ℃范围内,如灰绿青霉(Aspergillus glaucus)和罗尔夫青霉(Penicillium rolfsii)CCMB 714发酵产单宁酶的最适温度分别为40 ℃和30 ℃。不同野生型微生物发酵产单宁酶的最适pH不同,大多数野生型微生物发酵产单宁酶的最适pH为酸性至中性范围。野生型丝状真菌发酵产单宁酶的最适pH则偏酸性,如黑曲霉AUMC 5614和泡盛曲霉(Aspergillus awamori)BTMFW 032发酵产酶的最适pH为4.0和5.0。不同类型的野生型微生物的产单宁酶水平存在差异,野生型细菌和野生型酵母菌的产酶水平较低,而野生型丝状真菌产酶水平较高。地丝菌属酵母和肺炎克雷伯菌(Klebsiella pneumoniae)YL-1经发酵优化后产酶水平分别为4.4 U/mL和0.8 U/mL。黑曲霉AUMC 5614和泡盛曲霉BTMFW 032经发酵优化后产酶水平分别为11.6 U/mL和26.2 U/mL。青霉属液体发酵产单宁酶水平通常在5~40 U/mL之间。经发酵优化后,拜赖青霉CAU46-1产单宁酶水平达28.7 U/mL,高于皮壳青霉AN3和罗尔夫青霉CCMB 714发酵优化后的产单宁酶水平,分别为8.4 U/mL和25.6 U/mL,仅次于产紫青霉(Penicillium purpurogenum)产单宁酶水平(40.0 U/mL)。
2.3 单宁酶PbTanA的纯化
经硫酸铵沉淀和阴离子交换层析纯化得到单宁酶PbTanA的电泳级纯酶,比酶活力为34.2 U/mg,酶活力回收率为19%,纯化倍数为8.9(表1)。SDS-PAGE和酶谱对照确定其具有单宁酶活性,该酶呈单一蛋白条带,分子量为79.0 kDa(图4)。
表 1拜赖青霉CAU46-1单宁酶纯化
Table 1.Purification summary of tannase from Penicillium bilaiae CAU46-1
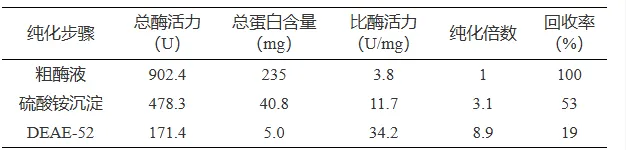
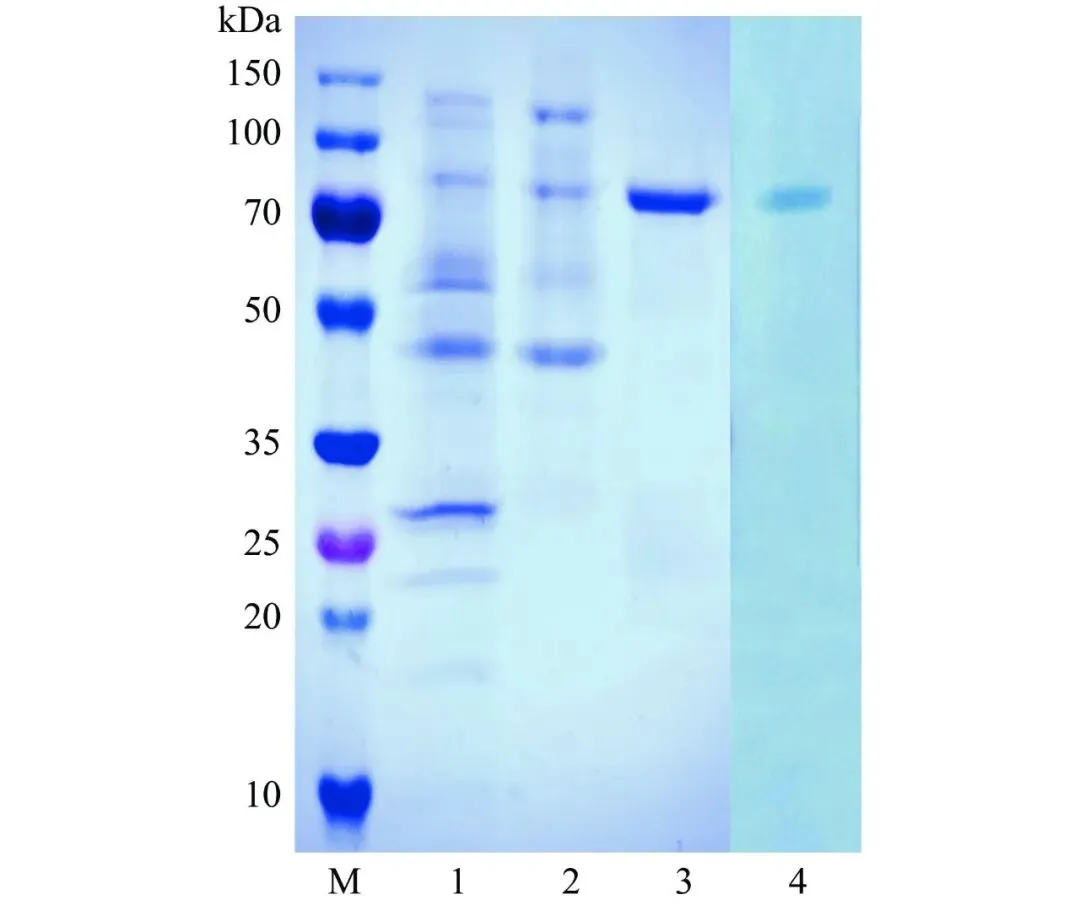
图 4拜赖青霉CAU46-1单宁酶PbTanA的纯化和酶谱分析
Figure 4.Purification and zymogram analysis of tannase (PbTanA) from Penicillium bilaiae CAU46-1
注:M:Marker;1:粗酶液;2:80%硫酸铵沉淀后;3:DEAE柱纯化后;4:酶谱分析。
不同来源单宁酶的分子量存在显著差异,细菌来源单宁酶一般分子量较小(31~90 kDa),而真菌来源单宁酶通常在45~320 kDa范围内。PbTanA的分子量高于地丝菌属酵母来源单宁酶(63 kDa)和黑曲霉NL112来源单宁酶(45 kDa),但低于黑曲霉LCF 8来源单宁酶(168 kDa)和米曲霉(Aspergillus oryzae)来源单宁酶(106 kDa)。
2.4 单宁酶PbTanA的酶学性质
PbTanA的最适pH为5.0(图5A),在pH2.0~7.0下处理30 min仍能保留80%以上的酶活力(图5B)。PbTanA的最适温度为60 ℃(图5C),在45 ℃以下处理30 min能够保留90%以上的酶活力,在50 ℃下处理30 mim仍保留70%以上的酶活力,具有良好的热稳定性(图5D)。不同金属离子及化合物对PbTanA酶活力影响不同(表2),金属离子Ag+、Cu2+对PbTanA的酶活力有明显抑制作用,酶活力分别下降至66.5%和61.4%;Co2+、Mn2+和Fe2+对酶活力有明显促进作用,酶活力分别提高至116.5%、112.5%和111.5%。EDTA处理对PbTanA酶活力影响较小,而SDS和β-巯基乙醇几乎完全抑制PbTanA的活性。
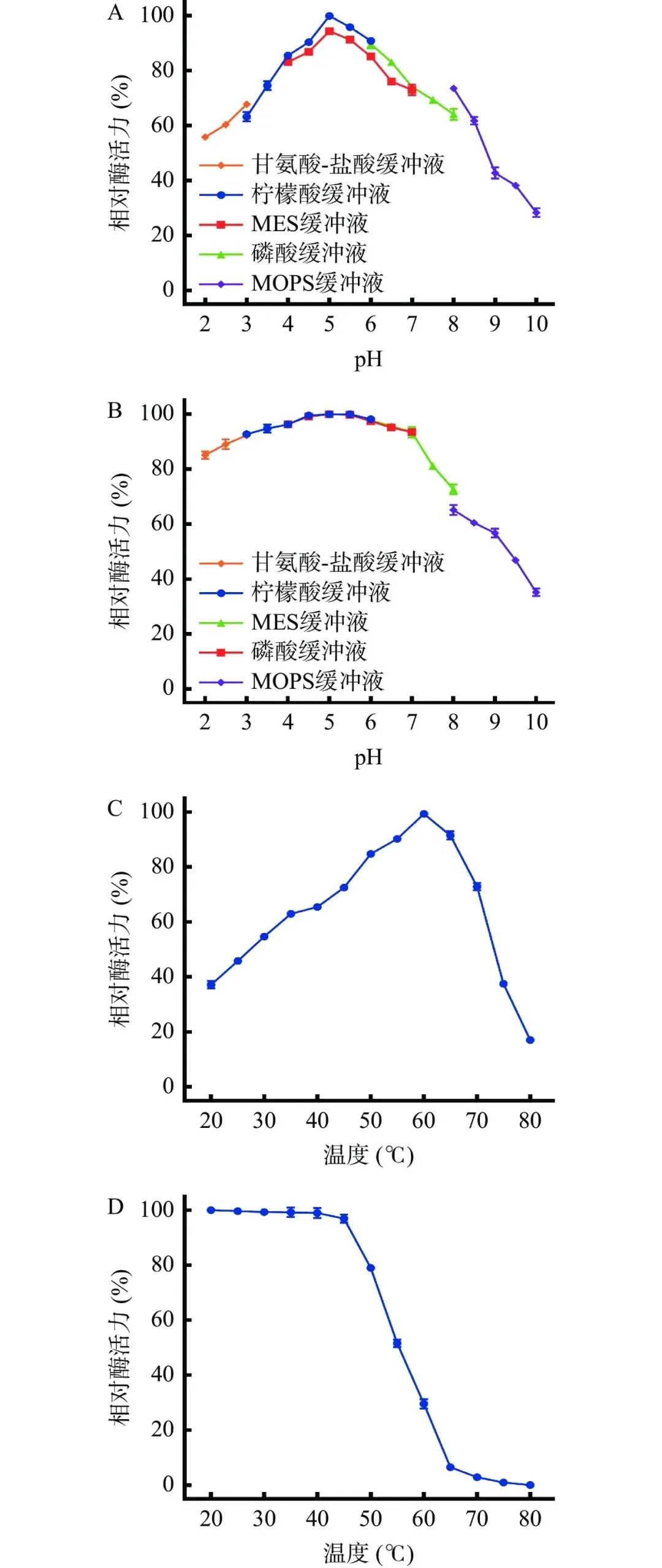
图 5PbtanA的最适pH(A)、pH稳定性(B)、最适温度(C)和温度稳定性(D)
Figure 5.Optimal pH (A), pH stability (B), optimal temperature (C), and temperature stability (D) of PbtanA
表 2金属离子和化合物对单宁酶PbtanA酶活力的影响
Table 2.Effects of metal ions and compounds on the enzymatic activity of PbtanA
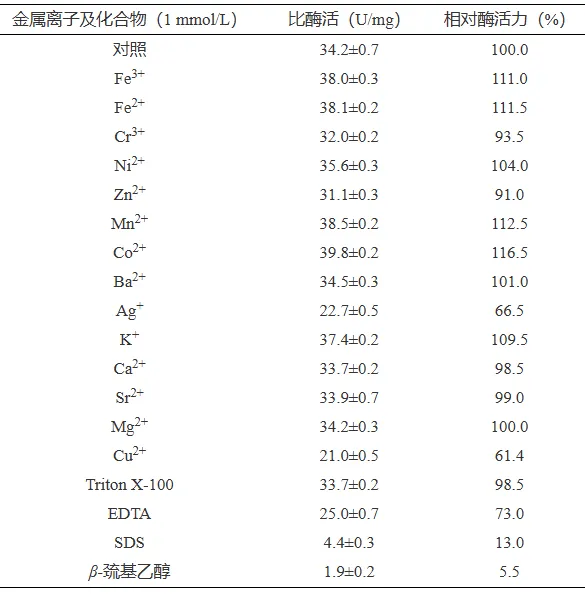
真菌来源单宁酶通常在酸性条件下具有较高的催化活性,如黑绿青霉EZ-ZH190和黑曲霉NL112来源的单宁酶,最适pH分别为5.5和5.0。PbTanA在酸性环境下具有良好的稳定性,在pH3.0下处理30 min仍保留80%以上的酶活力,优于黑绿青霉EZ-ZH190来源单宁酶(在pH3.5下处理30 min残留酶活力小于35%)和红绶曲霉(Aspergillus nomius)GWA5来源单宁酶(在pH4.0下处理30 min残留酶活力小于50%)。单宁酶PbTanA最适温度为60 ℃,明显高于地丝菌属酵母来源单宁酶(30 ℃)和青绿曲霉(Aspergillus glaucus)来源单宁酶(40 ℃)。此外,单宁酶PbTanA具有良好的热稳定性,高于黄曲霉TF-8来源的单宁酶(在50 ℃下处理30 mim,残留酶活力小于40%),但低于黑叶短梗霉(Aureobasidium melanogenum)T9来源单宁酶(55 ℃下处理12 h,能保留60%以上的酶活力)。不同金属离子对单宁酶的影响不同,如Mn2+可以抑制路黑曲霉ATCC 16620来源单宁酶的酶活力,却促进单宁酶PbTanA的酶活力。EDTA处理后,单宁酶PbTanA的残留酶活力为73%,表明EDTA抑制对PbTanA产生了一定的抑制作用,这与罗尔夫青霉CCMB 714来源单宁酶的酶学性质一致。SDS和β-巯基乙醇显著抑制PbTanA的酶活力,表明该酶的催化功能依赖于分子内二硫键的稳定。PbTanA具有良好的耐酸性和热稳定性,且对多种金属离子具有良好的耐受性,具备良好的应用潜力。
2.5 单宁酶的水解特性和水解单宁酸制备没食子酸
单宁酶PbTanA水解单宁酸历程如图6所示。PbTanA能够高效水解单宁酸,随着反应进行,产物没食子酸的生成量逐渐增多,以10、20和30 g/L单宁酸为底物,利用PbTanA反应8 h后没食子酸的浓度分别为9.3 g/L、18.4 g/L和24.5 g/L,得率分别为93.1%、91.8%、81.7%(图7)。
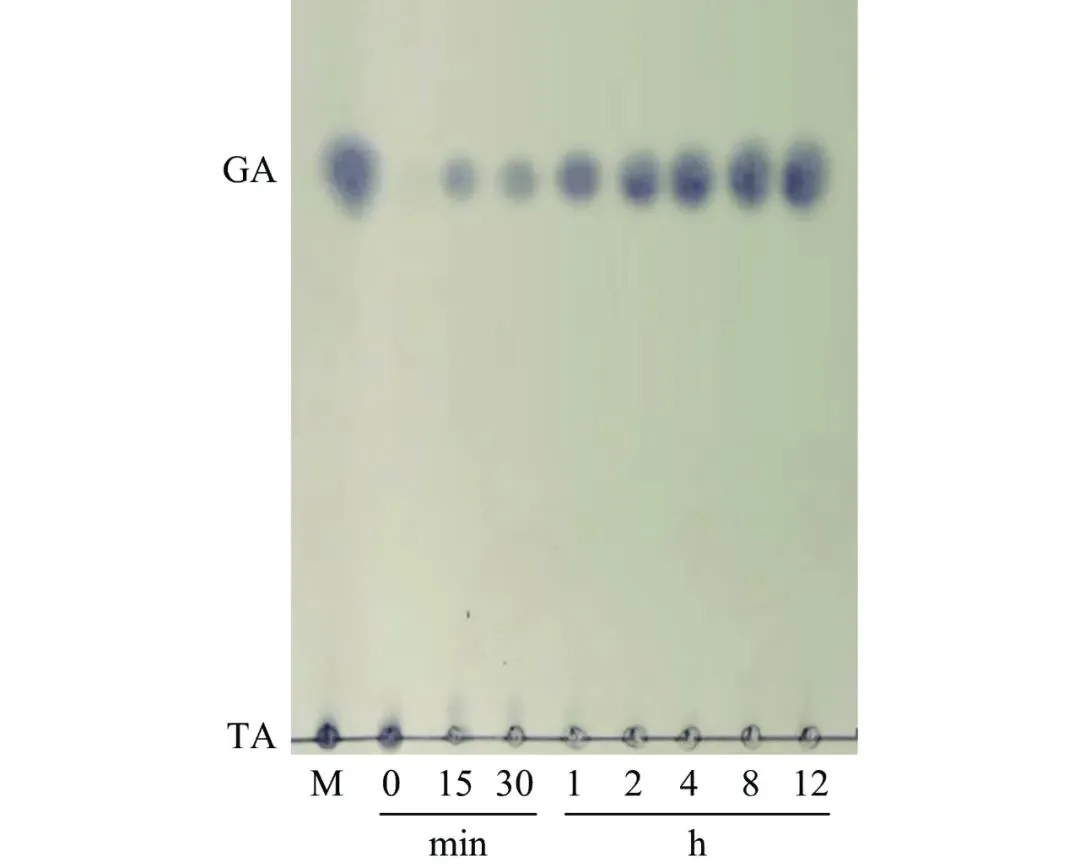
图 6单宁酶PbtanA水解单宁酸的TLC图
Figure 6.TLC for hydrolysis of tannic acid by PbTanA
注:M:标准品;GA:没食子酸;TA:单宁酸。
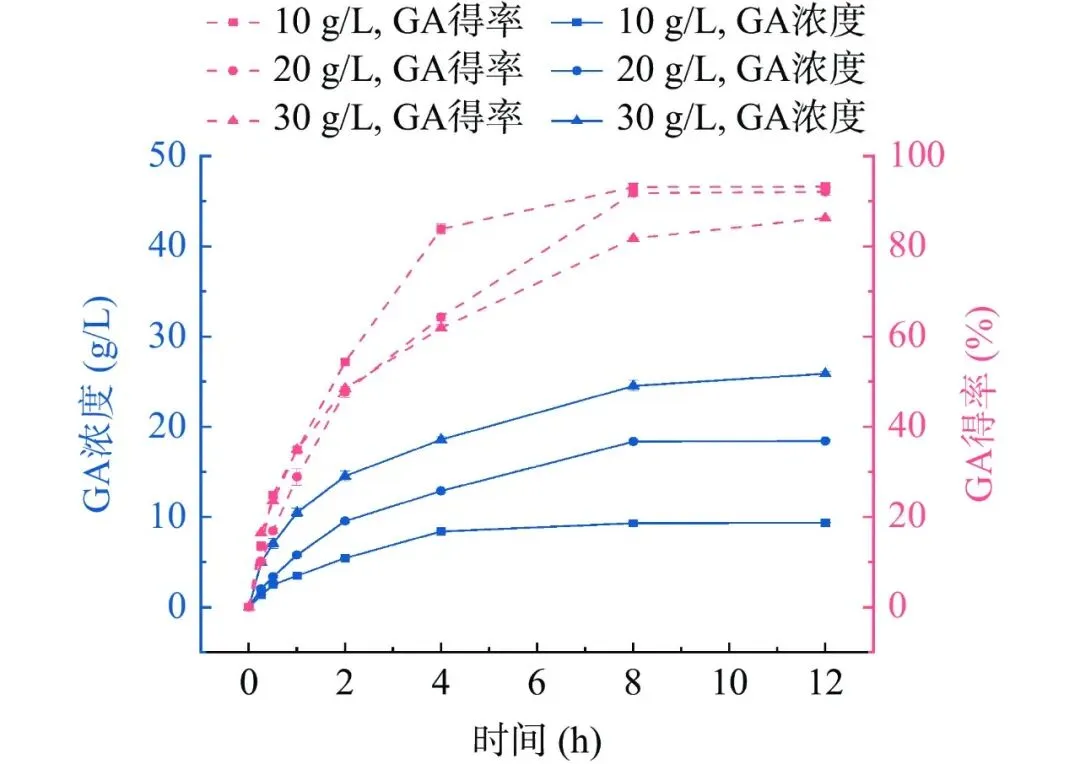
图 7单宁酶PbtanA水解不同浓度单宁酸生成没食子酸
Figure 7.Hydrolysis of gallic acid production from various concentrations of tannic acid by PbtanA
注:GA:没食子酸;TA:单宁酸;图例10、20、30g/L为TA浓度。
酶法制备没食子酸中,单宁酶作为关键催化剂,其水解特性和催化效率对生产效益具有决定性影响。球形芽孢杆菌(Bacillus sphaericus)来源单宁酶以50 g/L单宁酸为底物反应24 h,最终没食子酸得率为90.8%。汉森隐球菌来源单宁酶以20 g/L单宁酸为底物反应10 h,最终没食子酸得率91.6%。PbTanA水解20 g/L单宁酸底物8 h,没食子酸得率为91.8%,具有更高的生成量和得率,且反应时间更短,在水解单宁酸制备没食子酸中表现出很大的应用潜力。


本研究从大曲中分离出一株高产单宁酶的拜赖青霉CAU46-1。确定了拜赖青霉CAU46-1液体发酵产单宁酶的最适发酵产酶条件,产单宁酶水平(28.7 U/mL),较优化前提高了3.3倍。两步纯化得到电泳级纯酶PbTanA,该单宁酶具有良好的耐酸性和热稳定性,能高效水解单宁酸生成没食子酸,水解20 g/L单宁酸底物8 h,没食子酸得率为91.8%,在水解单宁酸制备没食子酸中展现出很大的应用潜力。本研究充分利用大曲微生物资源,拓展了单宁酶来源,为酶法制备没食子酸提供了理论指导。后续研究将结合质谱分析和基因组学分析,鉴定单宁酶PbTanA的基因序列,并实现其高效异源表达。
引用本文:杨淼,马志远,李延啸,等. 大曲中拜赖青霉发酵产单宁酶条件优化、纯化及酶学性质分析[J]. 食品工业科技,2026,47(8):212−220. doi: 10.13386/j.issn1002-0306.2025030409.
Citation:YANG Miao, MA Zhiyuan, LI Yanxiao, et al. Optimization of Fermentation Conditions for Tannase Production by Penicillium bilaiae from Daqu, and Its Purification and Analysis of Enzymatic Properties[J]. Science and Technology of Food Industry, 2026, 47(8): 212−220. (in Chinese with English abstract). doi: 10.13386/j.issn1002-0306.2025030409.


通讯作者简介

闫巧娟,博士,教授,教育部新世纪优秀人才,北京食品营养与人类健康高精尖创新中心岗位科学家。1997年硕士毕业于中国农业大学食品学院后留校任教,2004年晋升副教授,2009年晋升教授。长期从事农产品生物转化方面的科学研究,主要在食品相关酶制剂及其水解产物、功能性低聚糖、天然功能成分和微生物产有机酸等方面开展工作。近十年来,在木聚糖酶、葡聚糖酶、半乳糖苷酶和甘露聚糖酶等的产酶菌株筛选、基因克隆表达、酶的结构解析和分子改造、酶的纯化和性质及水解特性方面开展深入研究,得到多个性能优良的新型酶。主持完成国家863计划、国家自然科学基金项目等20多项。发表学术论文150余篇,其中SCI收录论文100余篇;授权国家发明专利40余项。获国家科技进步二等奖3项和省部级奖励3项。
(以上信息来自中国农业大学官网)
或登录www.spgykj.com阅览全文。
诚邀莅临

热门话题
查看更多 →
《食品工业科技》特邀主编专栏征稿



群聊:食品工业科技作者群

温
馨
提
示

我刊正式组建微信作者群,为作者提供更多的学术与论文资讯,如需进群,请联系刘老师(微信:上方二维码,电话:87244117-8062)。

版权声明



