华中农业大学猪遗传育种团队以多组学解析猪肌纤维类型的空间调控逻辑
- 2026-05-25 19:25:04
猪肉品质为何会在生长早期就埋下分化的“伏笔”?在看似相似的骨骼肌中,不同品种猪为何会走上不同的肌纤维发育轨迹?这一问题不仅关乎动物发育生物学的基础认知,也直接影响肉品质形成与遗传改良。近日,华中农业大学猪遗传育种团队在国际期刊 Advanced Science 在线发表题为 Multi-omics Insights into the Mechanisms of Early Muscle Fiber Difference and Transformation between Lean-type and Chinese Indigenous Pigs 的研究论文。该研究围绕瘦肉型猪与中国地方脂肪型猪出生后早期肌纤维差异形成机制这一关键科学问题,整合 RNA-seq、ATAC-seq、ChIP-seq、WGS 和 Hi-C 等多组学数据,系统揭示了表观组与三维基因组协同调控猪肌纤维类型的分子机制。

骨骼肌并非由单一细胞类型简单堆积而成,而是由代谢特征和收缩性能各异的多种肌纤维共同构成的复杂组织。已有研究表明,出生后早期是肌纤维类型转化的关键窗口,不同猪种在这一阶段的肌肉发育节律并不相同。然而,这种品种间差异究竟如何在染色质状态、顺式调控元件乃至空间基因组层面被建立起来,长期以来仍缺乏系统认知。
为回答这一问题,研究团队以 2 周龄瘦肉型猪(大白猪、杜洛克)和中国地方脂肪型猪(监利猪、莱芜猪)背最长肌为研究对象,首先从组织学和分子表型层面刻画了两类猪的早期差异图景。结果发现,在这一发育阶段,瘦肉型猪背最长肌的肌纤维平均直径和横截面积更小(图1A、B),但慢肌/氧化型相关特征更加突出,表现为更高比例的 MyHC1 阳性肌纤维,以及更高水平的 MyHC1、MyHC2a、SDH 和 MDH 等氧化代谢相关指标;相比之下,2周龄中国地方脂肪型猪背最长肌则呈现出更明显的 MyHC2b、MyHC2x 和 LDH 等糖酵解型特征(图1C、D)。这表明,在出生后不久,不同猪种便已沿着不同的肌纤维类型启动了各自的发育路径。
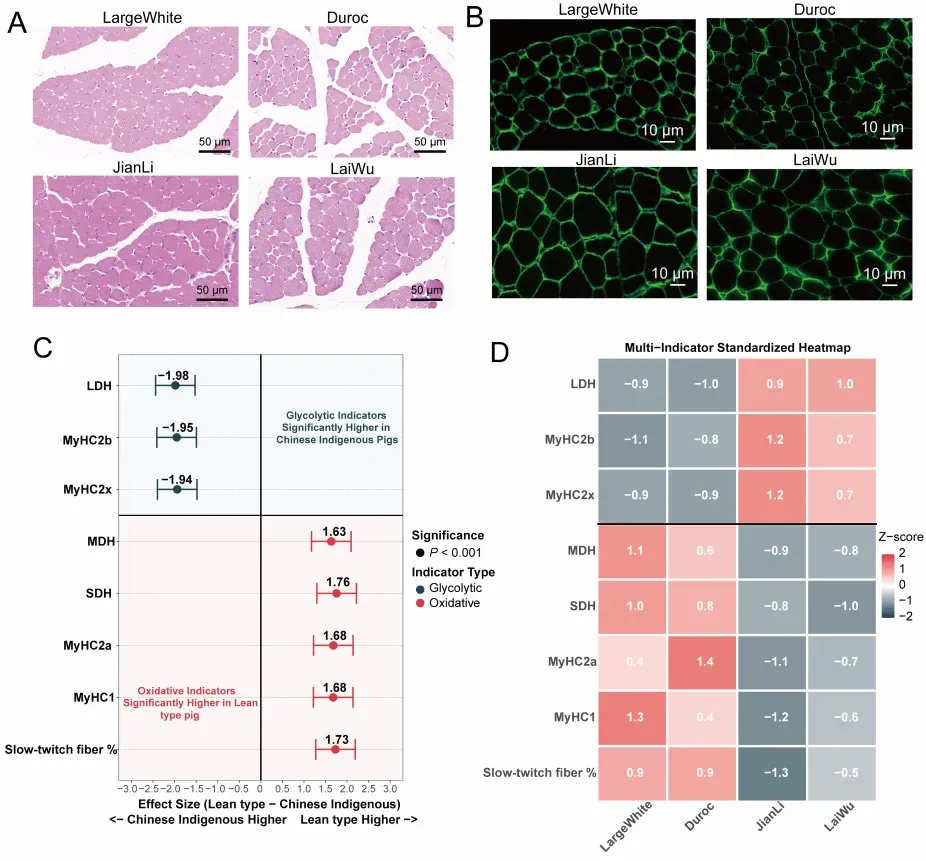
图1. 两周龄瘦肉型猪和中国地方脂肪型猪背最长肌的组织学及混合模型分析
但真正值得追问的是,肌纤维类型差异只是少数基因表达“高一点、低一点”的结果吗?研究团队进一步通过多组学整合分析发现,答案远比单一转录变化更为复杂。研究构建了猪早期骨骼肌的多层级调控图谱,发现瘦肉型猪与中国地方脂肪型猪之间不仅存在基因表达差异,还伴随着染色质开放性、增强子景观、启动子活性以及更高层级染色质结构的系统性分化(图2A)。尤其值得关注的是,三维基因组分析显示,两类猪之间存在 A/B compartment 转换、TAD 动态变化以及启动子-增强子互作网络重塑,提示肌纤维差异的形成并非仅发生在线性 DNA 序列层面,而是与染色质空间折叠方式密切相关(图2B、C)。
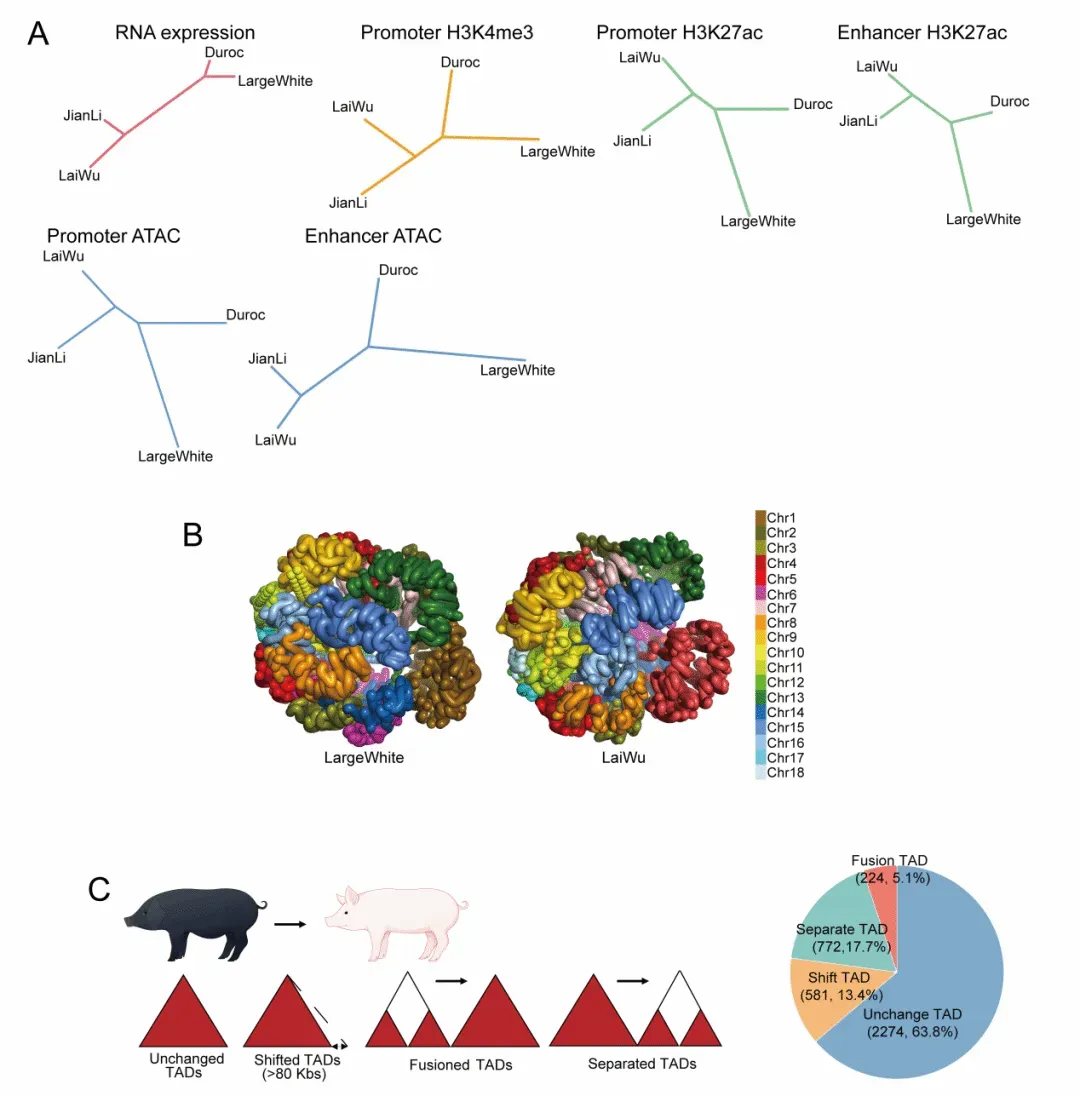
图2.两周龄瘦肉型猪与中国地方脂肪型猪背最长肌表观组及三维基因组学分析
在一系列关键发现中,研究团队鉴定到一个位于 PPP3CB 基因上游的瘦肉型猪特异性超级增强子(图3A)。通过体内外功能扰动实验发现,该超级增强子可通过招募转录因子 MEF2C,与 PPP3CB 启动子形成染色质互作,从而激活 PPP3CB 表达;随后,PPP3CB 促进NFATC1 去磷酸化,从而提升 MEF2C 表达,增强氧化代谢相关酶 SDH 和 MDH 活性,推动氧化型肌纤维形成。也就是说,PPP3CB 与 MEF2C 并非单向调控,而是形成了一个相互促进的正反馈调控环路:一方面,MEF2C 促进 PPP3CB 表达;另一方面,PPP3CB 又反向增强 MEF2C 表达,从而驱动氧化型肌纤维形成(图3B)。该研究揭示的 PPP3CB-MEF2C 正反馈调控环路,将“表观重塑-空间互作-基因激活-肌纤维类型”有机串联,加深了对猪肌纤维类型调控机制的认识,也为通过基因组和表观组策略改良肉品质提供了新的理论基础和分子靶点。
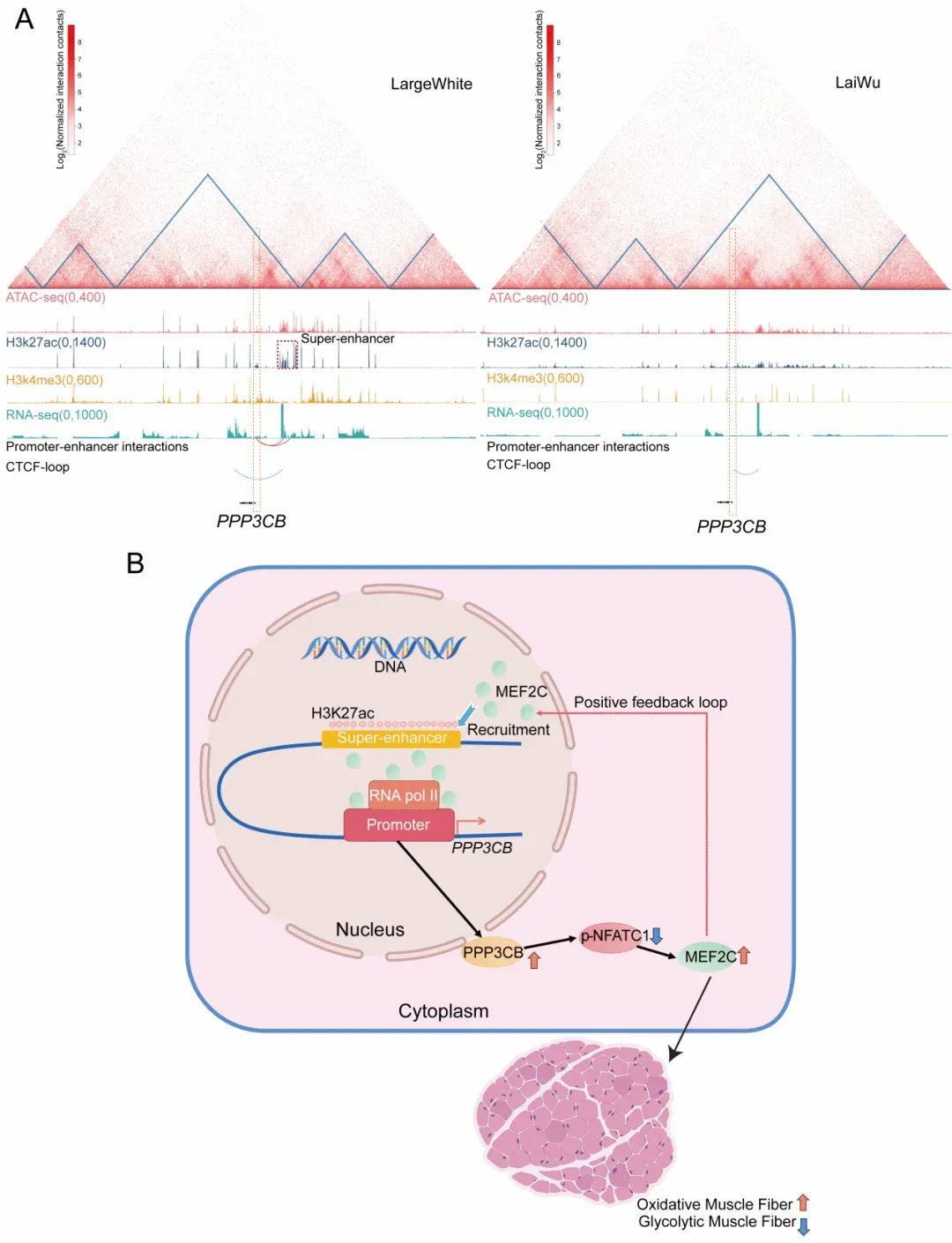
图3. 超级增强子-启动子互作通过PPP3CB-MEF2C 正反馈环路调控肌纤维类型
华中农业大学动物科学技术学院动物医学院徐德全教授和朱猛进教授为论文通讯作者,博士研究生郑帅龙和毕业硕士研究生吴海南为论文共同第一作者。该研究得到了国家重点研发计划等项目资助。
论文链接:https://advanced.onlinelibrary.wiley.com/doi/full/10.1002/advs.202523959
来源:南湖新闻网
畜牧人智库—不仅能找资料,还能与知识对话
扩展阅读
往期专题回顾




随机文章
-
 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
- 想拿中字头农业公司当门面,又怕被“原件不全”坑了?我们只做能验原件的硬通货!
- 黑龙江农业经济职业学院篇 | 2024—2025学年度黑龙江省国家奖学金获奖学生风采系列展示
- 【食品农业方向丨录用快丨早鸟优惠】2026年可持续农业、食品科技与营养健康国际会议(ICSAFTNH 2026)
- 五一华南农业大学继续教育学院放假通知!
- 校地协同强赋能 培育新农促振兴——赣州农业学校社会服务组培训工作再谱新篇
- 县农业农村局举办秸秆离田现场会
- 区政协副主席张胜涛开展特色农业产业专题调研
- 农业装备与能源工程团委|夯实团务基础,筑牢青春堡垒——农业装备与能源工程学院开展团支书团务知识考试
- 【现代农业在土左】逛田园、摘鲜果!五一就来土左旗现代农业示范区
- 【供】法国公司提供可持续农业生物解决方案
