3.1 燕麦叶片响应BYDV感染的差异表达基因鉴定
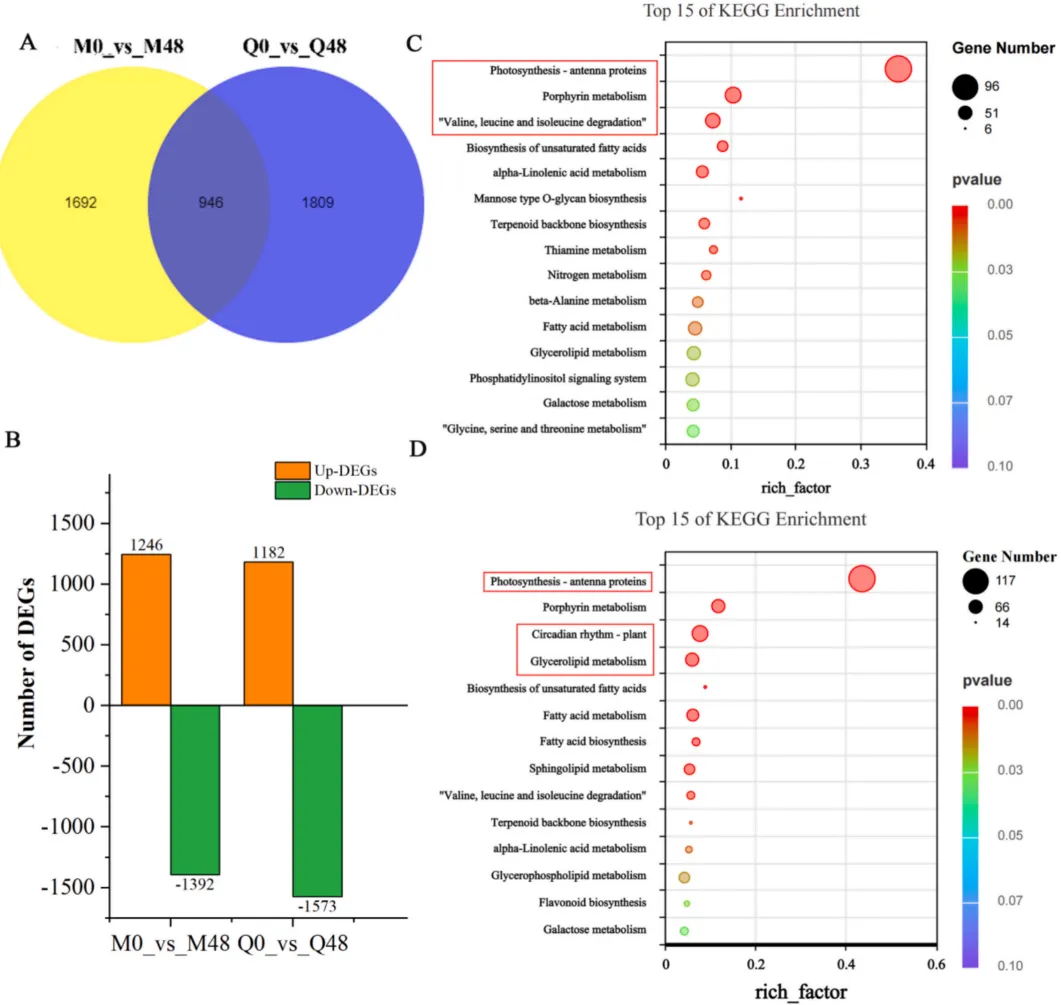
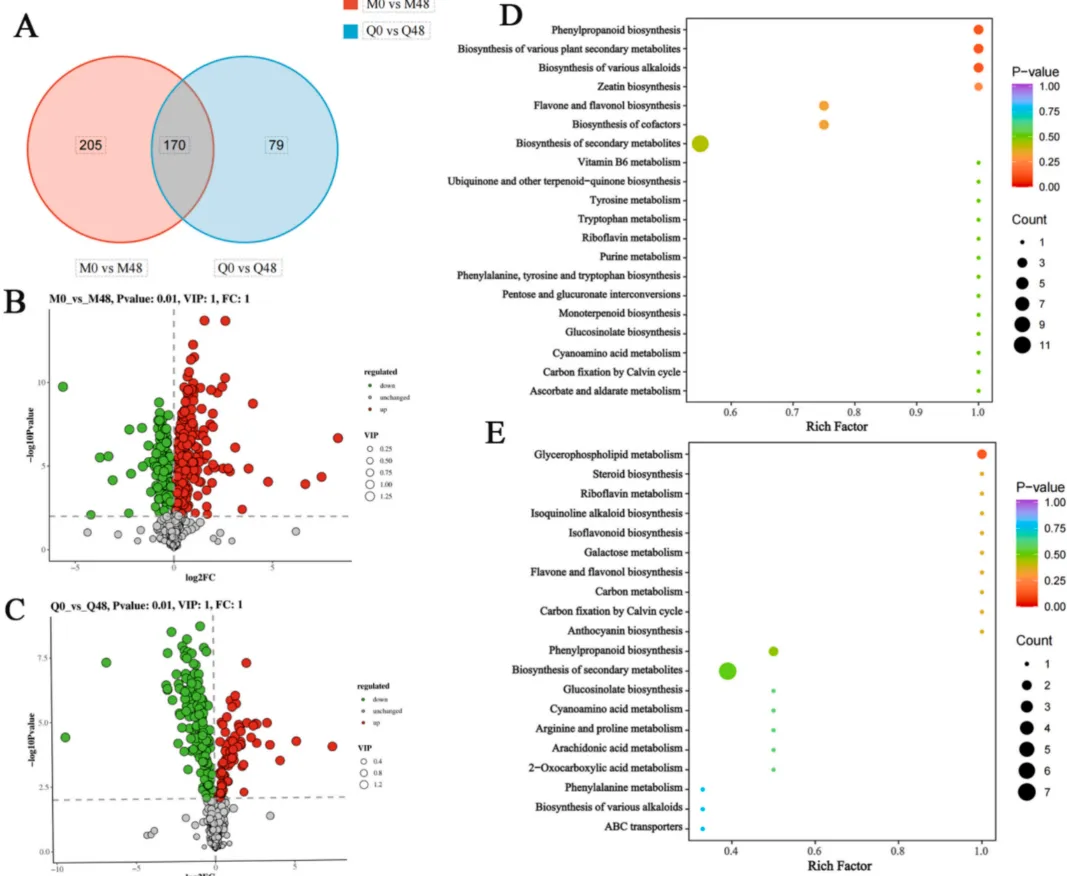
为评估不同抗性燕麦品种对BYDV的转录响应,研究对比了感染前后(0小时与48小时)的基因表达变化。结果表明,病毒感染在两个品种中均诱导了大量的基因表达重编程,但转录响应存在显著差异。抗性品种和易感品种在面对病毒胁迫时激活了截然不同的代谢通路,这暗示了其潜在的抗性机制分化(Fig.1)。
图1. 燕麦叶片感染BYDV后的转录组分析 两个比较组:M0_vs_M48(抗性品种MN10253)和Q0_vs_Q48(易感品种Qingyin 1)。(A) 韦恩图展示了两组间共有和特有的DEGs数量。(B) 与0小时相比,MN10253和Qingyin 1在感染后48小时鉴定出的上调和下调DEGs总数。(C, D) MN10253 (C) 和Qingyin 1 (D) 中DEGs的KEGG通路富集分析,突出了受BYDV感染影响的关键生物学途径。
3.2 加权基因共表达网络分析(WGCNA)
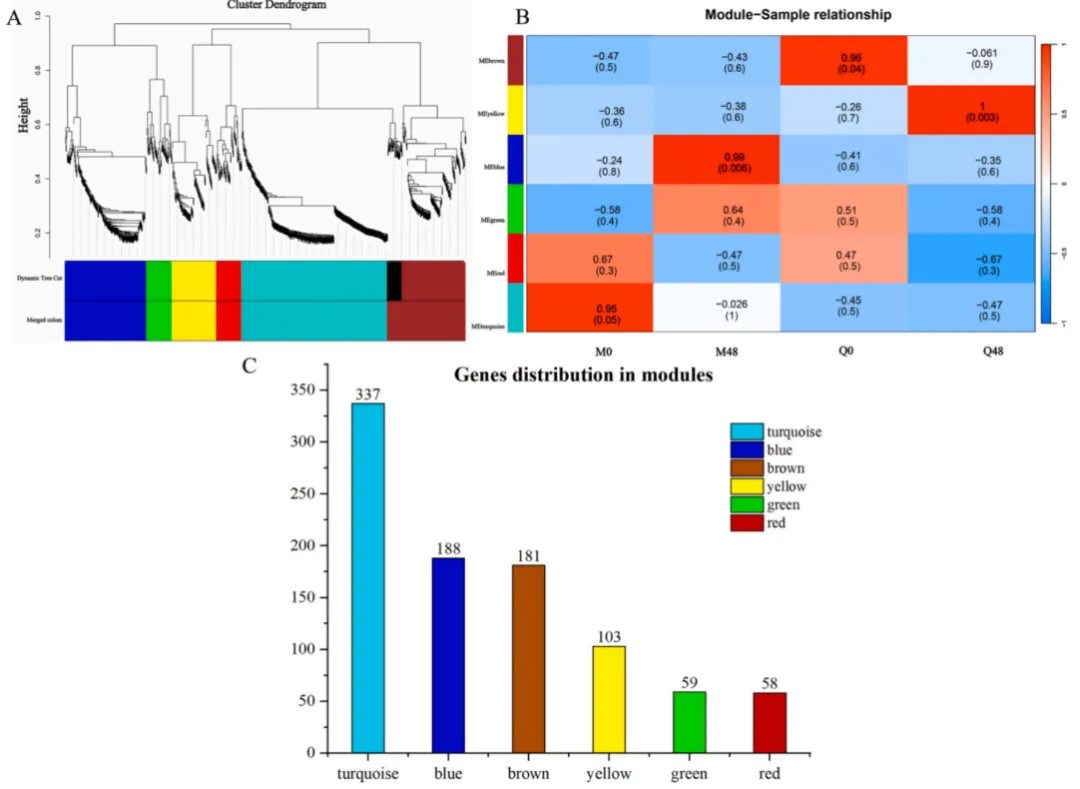
为深入探究响应BYDV感染的基因共表达模式,采用WGCNA方法对转录组数据进行了模块化分析。结果揭示了各共表达模块在不同时间点和不同品种间呈现出特异性的表达趋势。这种高度的模块-样本相关性表明,每个品种在BYDV感染的不同阶段均表现出独特的基因共表达网络,为理解抗性与易感性背后的动态转录调控提供了重要线索(Fig.2)。
图2. 基因共表达聚类与模块-样本相关性分析 (A) 所有样本中共表达基因的层次聚类,形成六个不同的模块。(B) 热图展示了每个模块与样本之间的相关性,说明了不同时间点和品种间基因表达模式的差异。(C) 每个共表达模块内基因数量的分布。
3.3 燕麦叶片响应BYDV感染的代谢组学分析
通过对感染BYDV前后燕麦叶片的代谢组学分析,揭示了抗性与易感品种在代谢水平上的显著差异。抗性品种MN10253在感染后积累了大量上调的代谢物,表明其具有更活跃的与抗病性相关的代谢响应。KEGG富集分析进一步证实,两个品种在应对病毒胁迫时激活了完全不同的代谢网络,抗性品种更倾向于次生代谢防御(Fig.3)。
图3. MN10253和Qingyin 1燕麦叶片响应BYDV感染的代谢组学概貌 (A) 韦恩图展示了MN10253 (M0_vs_M48) 和Qingyin 1 (Q0_vs_Q48) 中差异表达代谢物 (DEMs) 的分布。(B, C) 火山图说明了MN10253 (B) 和Qingyin 1 (C) 中上调和下调的DEMs。(D, E) MN10253 (D) 和Qingyin 1 (E) 中DEMs的KEGG通路富集分析。
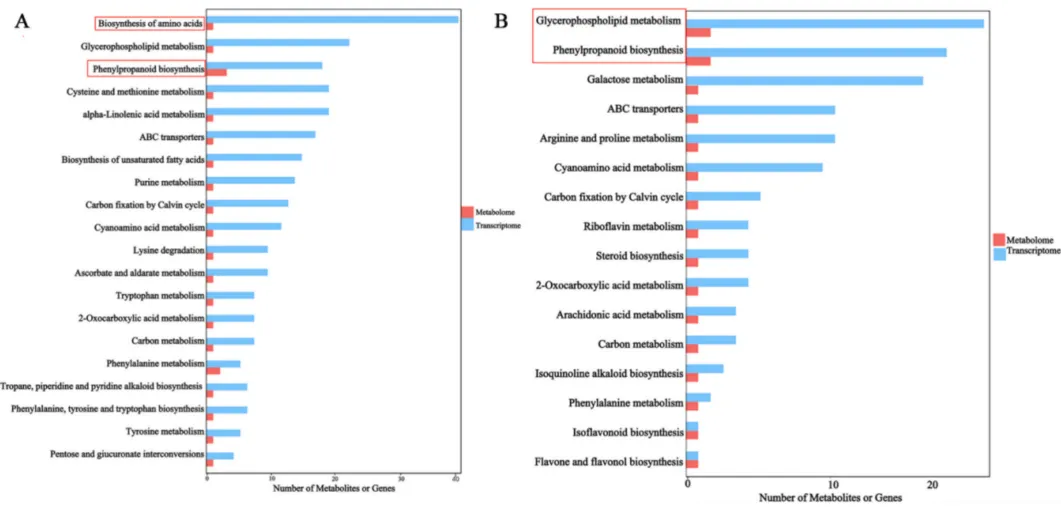
3.4 DEGs与DEMs的联合富集分析
综合转录组和代谢组数据的KEGG通路联合富集分析,精准定位了燕麦响应BYDV感染的核心代谢途径。结果表明,苯丙烷和氨基酸代谢途径在燕麦的防御反应中发挥着至关重要的作用,特别是这些途径的协同激活可能是增强抗性品种对BYDV抵御能力的关键分子基础(Fig.4)。
图4. MN10253 (A) 和 Qingyin 1 (B) 中DEGs和DEMs的KEGG通路联合富集分析 使用超几何检验计算富集显著性。红色箭头指示富集的代谢物,蓝色箭头指示苯丙烷生物合成 (ko00940)、氨基酸生物合成 (ko01230) 和甘油磷脂代谢 (ko00564) 途径中富集的基因。
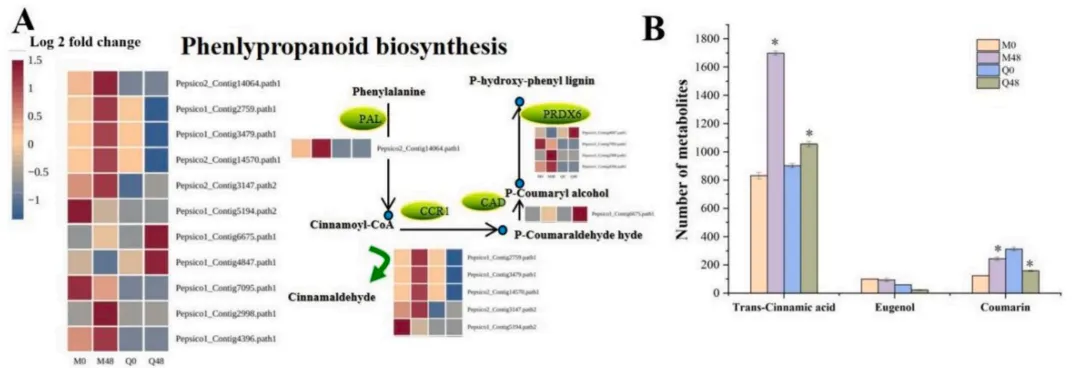
3.5 BYDV感染后的苯丙烷代谢途径
KEGG通路分析深入揭示了苯丙烷生物合成途径在两品种间的差异调控。抗性品种通过显著上调该途径的关键酶基因,促进了相关防御性次生代谢物的合成。特别是香豆素在抗性与易感品种间截然相反的积累模式,强烈暗示其可能作为燕麦抵御BYDV感染的关键防御代谢物发挥作用(Fig.5)。
图5. BYDV感染后MN10253和Qingyin 1中苯丙烷生物合成途径的基因表达 (A) 和代谢物 (B) 变化 红色突出的基因表示上调,蓝色表示下调。色标对应于log2倍数变化值。星号表示与同一基因型对应的未接种0小时对照相比存在显著差异 (p < 0.05)。
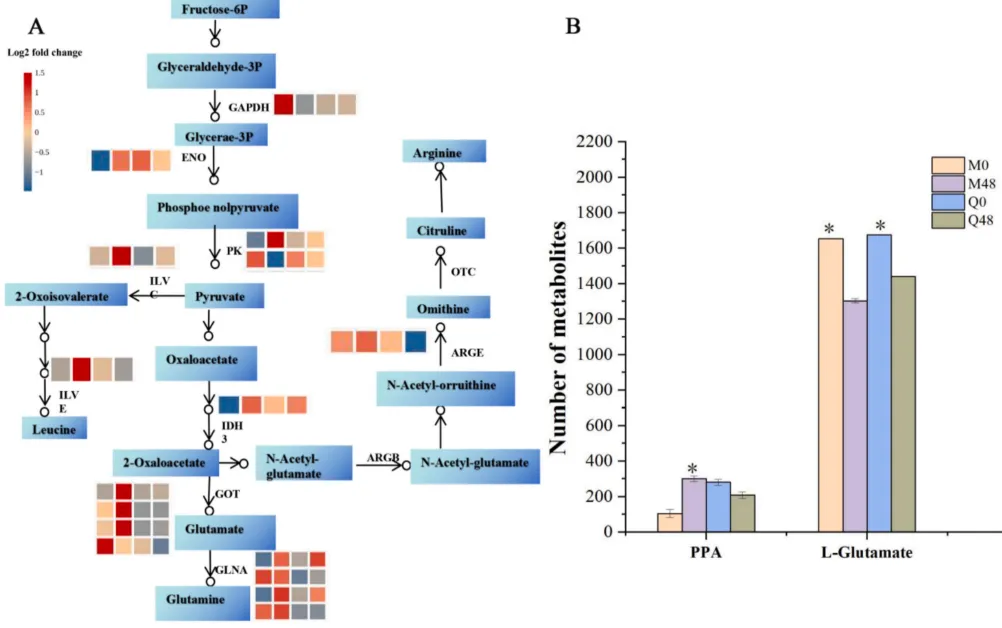
3.6 BYDV感染后氨基酸生物合成途径的变化
基于氨基酸代谢网络的综合分析表明,抗性品种在BYDV感染后能够更有效地重塑氨基酸生物合成途径。通过上调多种关键代谢酶的表达,不仅促进了特定氨基酸及其衍生物(如PPA)的积累,还揭示了氨基酸代谢在介导燕麦抗病毒防御响应中不可或缺的调节作用(Fig.6)。
图6. MN10253和Qingyin 1感染BYDV后氨基酸生物合成差异调控的通路图 红色突出的基因表示上调,蓝色表示下调。星号表示与同一基因型对应的未接种0小时对照相比存在显著差异 (p < 0.05)。
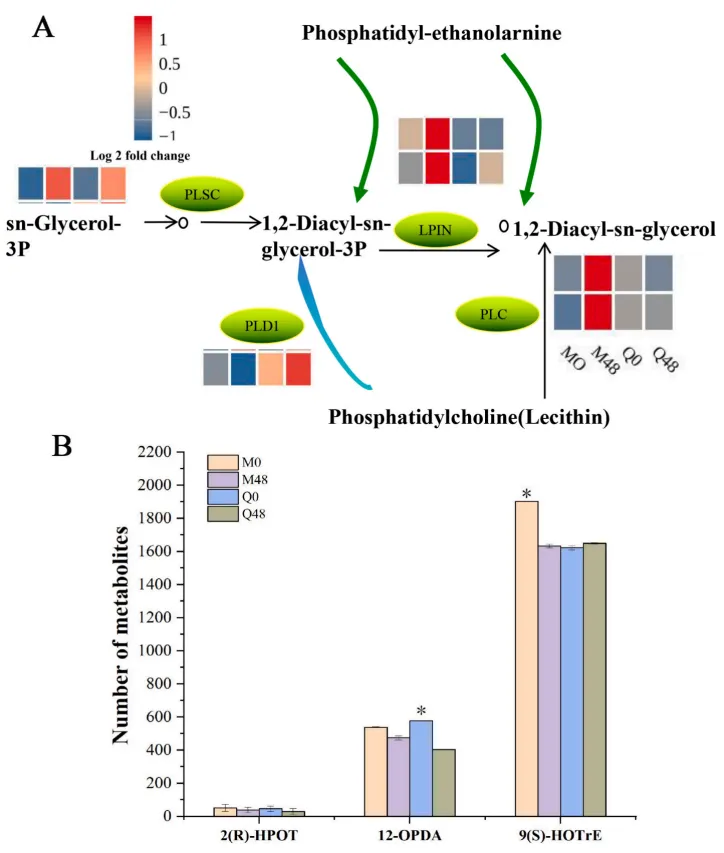
3.7 BYDV感染下的甘油磷脂代谢途径
甘油磷脂代谢途径的重编程是燕麦响应BYDV胁迫的重要环节。抗性品种通过特异性上调或下调特定的磷脂酶基因,精细调控了脂质代谢网络。同时,关键信号脂质分子的显著消耗,暗示了甘油磷脂代谢及其偶联的脂质信号传导机制可能积极参与了燕麦的抗病毒防御反应(Fig.7)。
图7. 与甘油磷脂代谢途径相关的DEGs 最大/最小log2 FPKM值设置为±2.0(红色表示上调,蓝色表示下调)。每条水平行代表一个DEG,垂直列分别代表两个基因型(MN10253和Qingyin 1)的两个时间点(0,48小时)。星号表示与同一基因型对应的未接种0小时对照相比存在显著差异 (p < 0.05)。
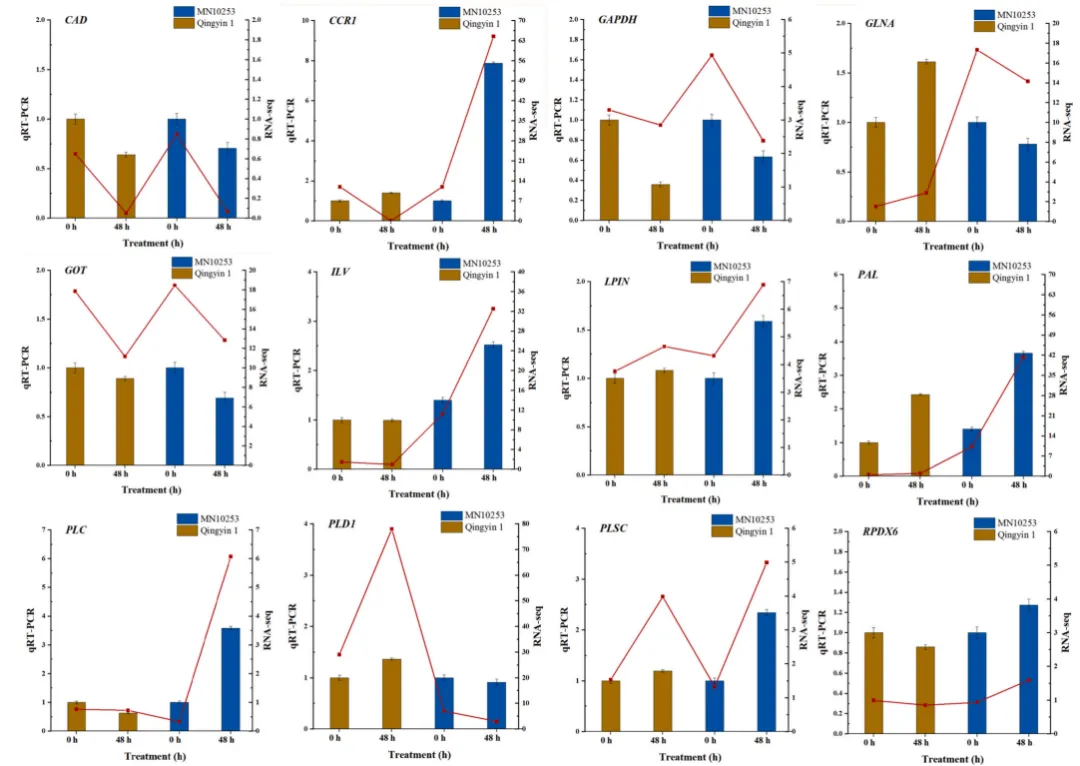
3.8 关键通路基因的qRT-PCR验证
为确保组学数据的准确性,采用qRT-PCR对关键代谢途径的核心基因进行了表达量验证。验证结果不仅证实了RNA-seq数据的稳健性,更进一步凸显了抗性与易感品种在面对BYDV-GPV感染时截然不同的转录响应模式,深刻反映了两者潜在防御机制的本质差异(Fig.8)。
图8. 接种BYDV-GPV后抗性 (MN10253) 和易感 (Qingyin 1) 燕麦品种关键通路中代表性基因的qRT-PCR验证 分析了0小时和接种后48小时苯丙烷生物合成 (CCR1, PAL, CAD)、氨基酸生物合成 (GAPDH, GLNA, GOT, ILV, RPDX6) 和甘油磷脂代谢 (PLSC, PLD1, LPIN, PLC) 途径中12个代表性基因的表达模式。折线图代表RNA-seq表达数据,而条形图显示qRT-PCR验证结果。