水淹胁迫对LS273和ZZ39茎和根形态生理的影响
研究发现,LS273和ZZ39在水淹胁迫下的生长表现存在显著差异,LS273相对于ZZ39表现出更高的根长、茎高和鲜重,根表面积、体积、尖端数量也有所增加。LS273在水淹胁迫下的茎高和根长分别比ZZ39高出142.65%和250.00%。这表明水淹胁迫显著影响水稻的生长,尤其是根部的生长,而LS273通过快速生长特别能够抵抗水淹带来的缺氧胁迫。
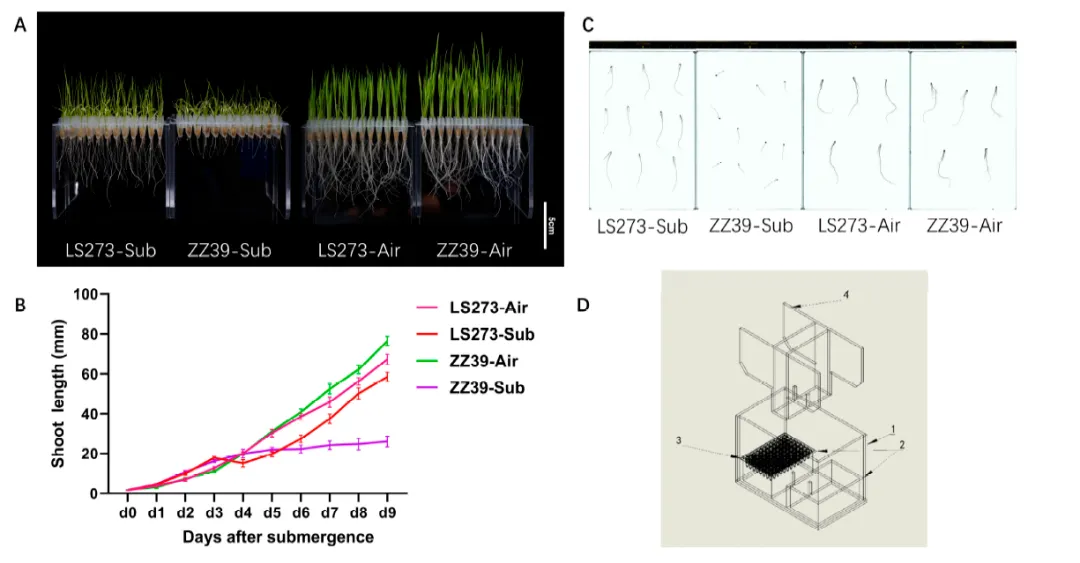
图1. 正常和水淹条件下LS273和ZZ39品种芽生长的形态特征。(A) LS273与ZZ39第9天表型差异(B) LS273与ZZ39幼苗高度生长动态。(C) ZZ39和LS273的根扫描图(分辨率300 ppi)。(D)水淹鉴定设备示意图:(1)淹水箱,(2)根阴箱,(3)开孔PCR板,(4)上压板。LS273- Sub:LS273淹没处理;ZZ39-Sub:LS273水淹处理;LS273- Air:LS273空白对照;ZZ39- Air:ZZ39空白对照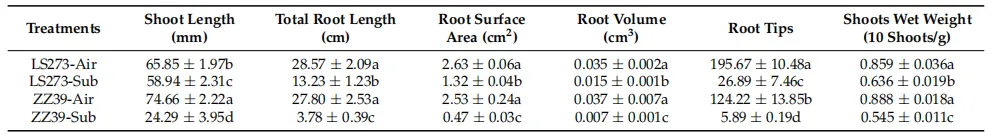
表1. 水淹胁迫影响不同抗性水稻的表型。同字母列差异无统计学意义(p < 0.05),单因素方差分析,随后进行HSD检验或其他检验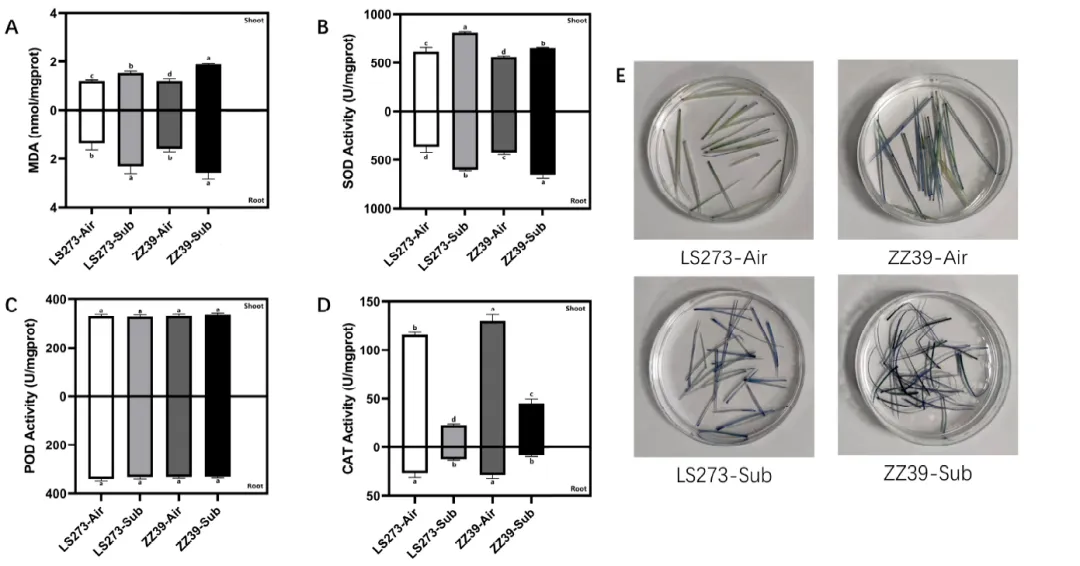
图2. 抗氧化酶及TB染色结果。(A)丙二醛(MDA)含量;(B) 超氧化物歧化酶(SOD)活性;(C)过氧化物酶(POD)活性;(D) 过氧化氢酶(CAT)活性;(E) TB染色结果。同字母列差异不显著,p < 0.05,单因素方差分析,其次是HSD差异等检验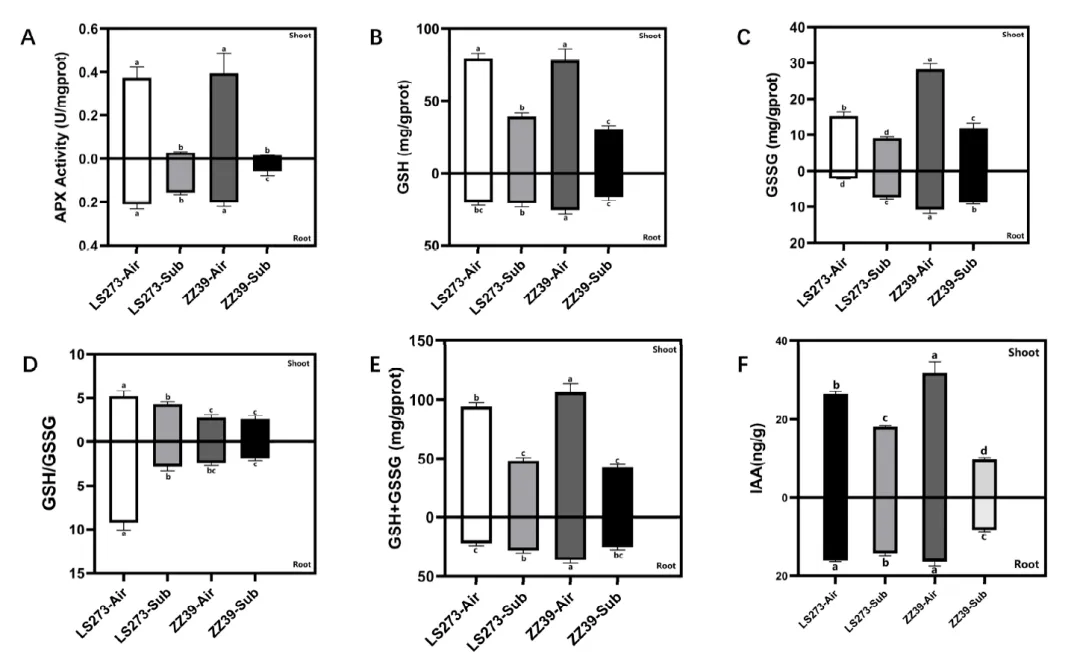
图3. 谷胱甘肽循环和生长激素测量。(A) APX活性;(B) GSH含量;(C) GSSG含量;(D)谷胱甘肽(GSSG);(E) GSH + GSSG含量;(F) IAA含量。同字母列差异不显著(p < 0.05),单因素方差分析,其次是HSD差异检验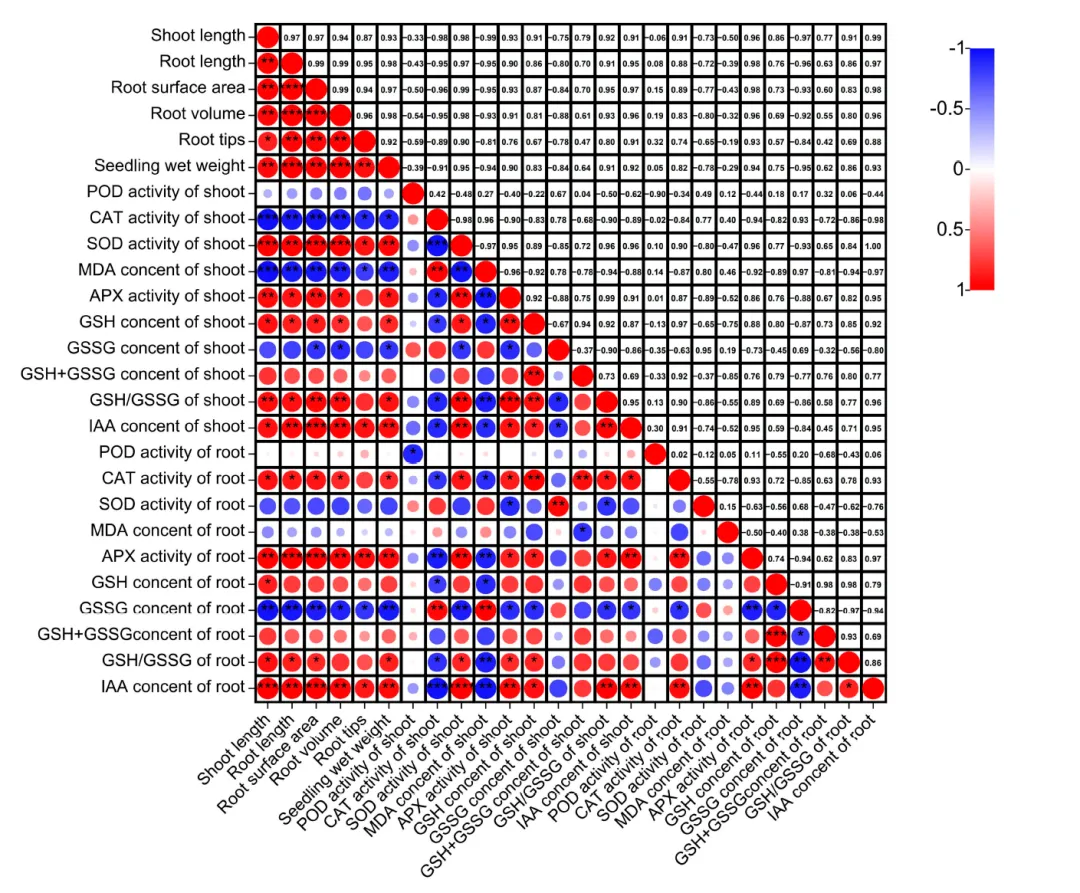
图4. 水淹胁迫下幼苗生长与生理指标的相关性分析。****:在0.0001概率水平上显著(p < 0.0001);***:在0.001概率水平上显著(p < 0.001);**:在0.01概率水平上显著(p < 0.01);*:在0.05概率水平上显著(p < 0.05)
RNA测序(RNA- seq)分析和鉴定差异表达基因(DEGs)
研究中提取了在水淹胁迫和空白对照条件下生长的LS273和ZZ39的茎和根的RNA,样本采集时间为播种后的第7天。RNA测序分析结果显示,平均每个样本产生了42.86–73.62百万条原始数据,92.47%为过滤数据,具有94.20%以上的高质量分数。基因表达程度通过每百万个片段映射的每千碱基转录本的碎片数(FPKM)进行测定,基因表达丰度则使用Cufflink软件进行分析。PCA主效应组件分析显示,不同处理的生物重复样本明显聚集,表明转录组分析结论的可信度。基因表达的变异中,PC1(65.6%)的最大变异源于组织差异,PC2(18.92%)的变异源于品种差异,表明不同组织之间的转录组变异明显。不同品种之间的差异最不重要,而不同组织之间的差异最显著。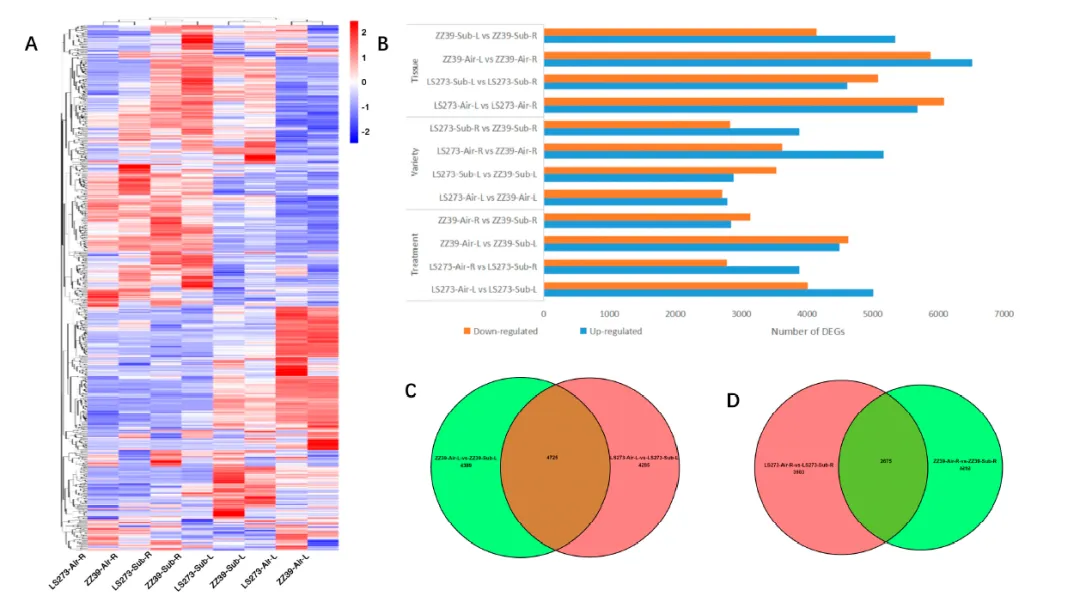
图5. 水淹条件下的DEGs。(A)处理间DEGs热图;(B)不同组织、品种和处理中DEGs的数量;(C)水淹条件下LS273和ZZ39幼苗的独有和共有DEGs;(D)水淹条件下LS273和ZZ39根系中独有和共有的DEGs
在LS273中有4285个独有的DEGs,ZZ39中有4389个独有的DEGs,共有4725个DEGs。为了了解这些特殊应答基因在水淹胁迫期间茎部的功能,还对DEGs进行了GO分析和KEGG分析。GO分析发现LS273茎部的NAD代谢过程(GO:0019674)和碳水化合物代谢过程(GO:0005975)受到严重影响。此外,氧化还原酶活性(GO:0016491)和铁离子结合(GO:0005506)在LS273中也高度富集。在ZZ39的茎部中,光合电子传递系统II(GO:0009772)、光系统II(GO:0009523)和光系统I(GO:0009522)受到主要影响。此外,丝氨酰-tRNA氨酰化(GO:0006434)在ZZ39中也高度富集。这两个品种的茎部参与了氧化还原过程(GO:0055114)和碳水化合物代谢过程(GO:0005975)。此外,脂肪酸生物合成过程(GO:0006633)、脂质代谢过程(GO:0006629)、铁离子结合(GO:0005506)、细胞壁(GO:0005618)和糖酵解过程(GO:0006096)也显著富集。KEGG通路分析显示,LS273主要在过氧化物酶体(dosa04146)、植物激素信号传导(dosa04075)、脂肪酸代谢(dosa01212)和黄酮类和类黄酮类生物合成过程(dosa00944)方面得到改善。ZZ39主要富集于次生代谢产物的生物合成(dosa01110)、氨酰-tRNA生物合成(dosa00970)以及哌啶和吡啶生物碱生物合成(dosa00960)。这两个品种的茎部富集于碳代谢(dosa01200)、次生代谢产物的生物合成(dosa01110)、代谢途径(dosa01100)和苯丙烷生物合成(dosa00940)。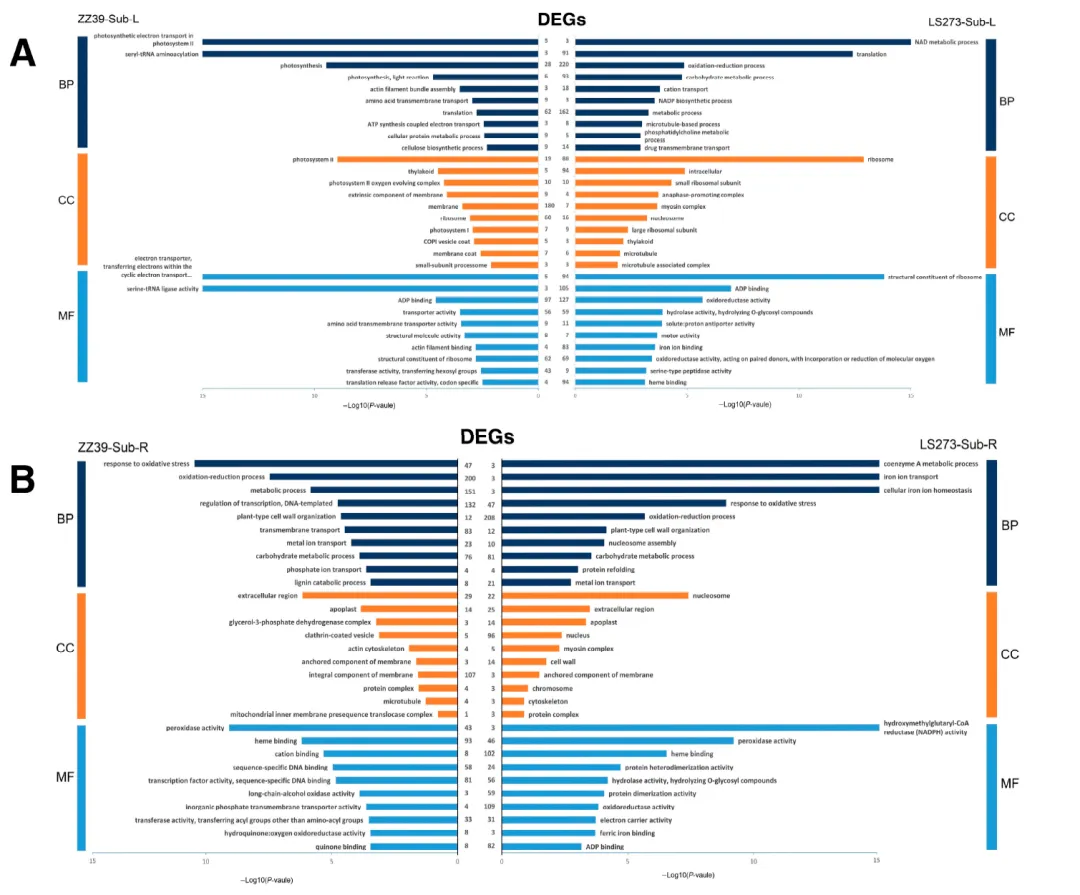
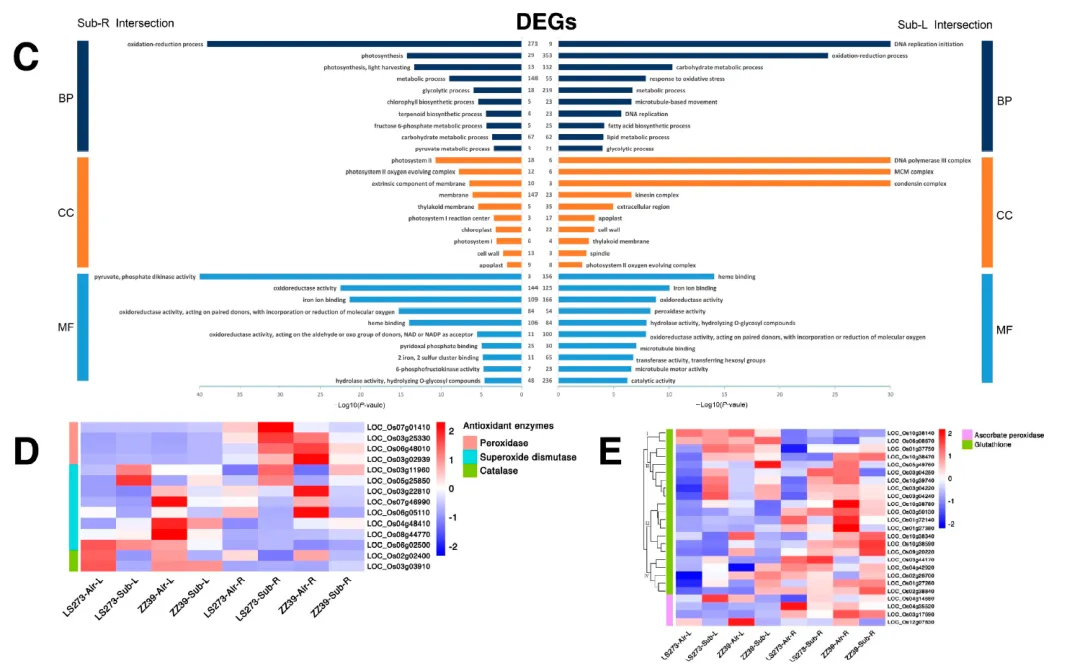
图6. 水淹胁迫下的富集和抗氧化系统下DEGs的表达。(A)芽部DEGs的富集结果;(B)根中DEGs的富集结果;(C)与根共享DEGs的富集结果;(D)抗氧化酶系统中DEGs的表达;(E)谷胱甘肽循环中DEGs的表达。BP:生物过程;CC:细胞成分;MF:分子功能;I、II、II、IV:聚类分析类型通过对SOD、CAT和POD等主要抗氧化酶的分析,KEGG分析表明水淹胁迫显著影响了水稻茎部的氧化还原系统。在LS273中,18个与抗氧化酶方法相关的DEGs被发现,其中一些基因在水淹胁迫后在茎和根中上调。与LS273相比,ZZ39在水淹胁迫后在茎部上调而在根部下调。不同种类的SOD在不同部位对水淹胁迫的反应中表达差异。此外,与CAT相关的基因在LS273的茎部表达高于根部。APX是植物活性氧代谢中最重要的抗氧化酶之一,特别是叶绿体中清除H2O2的关键酶。与APX相关的四个DEGs在不同品种和组织中表达不同。此外,通过AsA-GSH循环,谷胱甘肽可以清除增加的过氧化氢。在水淹胁迫后,与谷胱甘肽相关的21个DEGs被鉴定。这些基因在不同组织和品种中表达差异,并在水淹胁迫后显示出显著的表达变化。
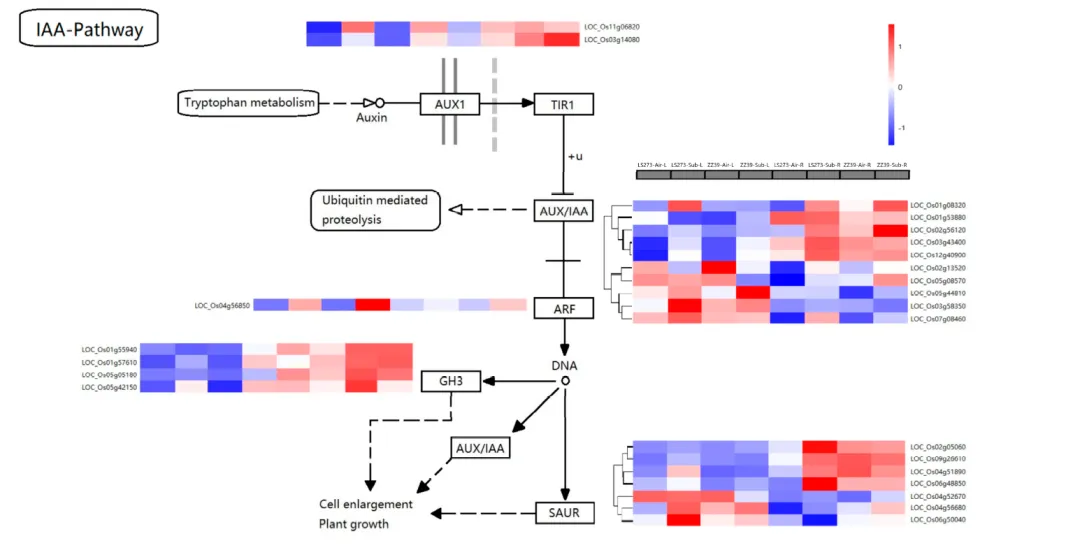
根据KEGG分析,植物激素信号传导通路在这一过程中得到显著丰富。通过差异基因筛选,发现了24个与生长素相关的DEGs。这些DEGs在生长素代谢中的作用得到了分析。在这些基因中,AUX1相关基因LOC_Os11g06820和LOC_Os03g1408在水淹胁迫下上调。生长素早期响应基因包括AUX/IAA、GH3和SAUR基因家族。这些基因在水淹胁迫下的表达存在差异。在AUX/IAA家族基因中,有十个被分类为四类。在水淹胁迫下,这些基因的表达在LS273和ZZ39的茎和根中有所不同。一些基因在茎部上调,而在根部下调。其他基因则在水淹胁迫下在茎和根中均上调。还有一些基因在水淹胁迫下在茎部下调而在根部上调。这些结果揭示了生长素在水稻对水淹胁迫的调节和抗性中的重要作用。研究选取了三个谷胱甘肽S-转移酶相关基因进行检测。在LS273的茎部,LOC_Os01g72140的表达显著上调,但在ZZ39的茎部下调;LOC_Os10g38540在LS273的根部显著下调,但在ZZ39的根部显著上调;LOC_Os03g04220在LS273的根部显著上调,但在ZZ39的根部显著下调。另外,LOC_Os03g17690(OsAPX1)在LS273中表达无差异,但在ZZ39中上调。尽管在ZZ39的茎部下调,LOC_Os12g07830(OsAPX5)在LS273的茎部上调。在LS273的茎和根中表达不同,在ZZ39的茎部上调的LOC_Os03g03910(OsCATC)。LOC_Os01g55940(OsGH3-2)在ZZ39中上调,在LS273中下调。在水淹胁迫后,LOC_Os05g42150(OsGH3-4)在茎部上调,在根部下调。SAUR基因家族的LOC_Os02g05060的表达在水淹胁迫后根部高于茎部。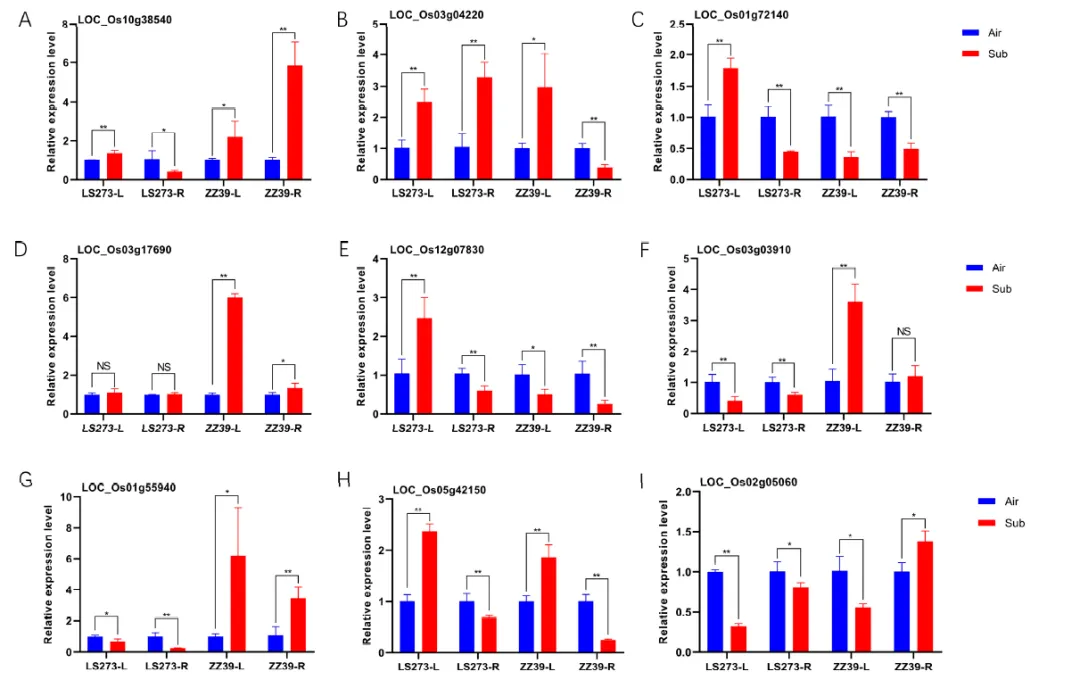
图8. 实时荧光定量PCR (q-PCR)测定品种不同部位抗氧化相关基因(A-F)。实时荧光定量PCR (q-PCR)测定不同品种(G-I)不同部位生长素相关基因。**:在0.01概率水平上显著(p < 0.01);*:在0.05概率水平上显著(p < 0.05);NS:无显著差异