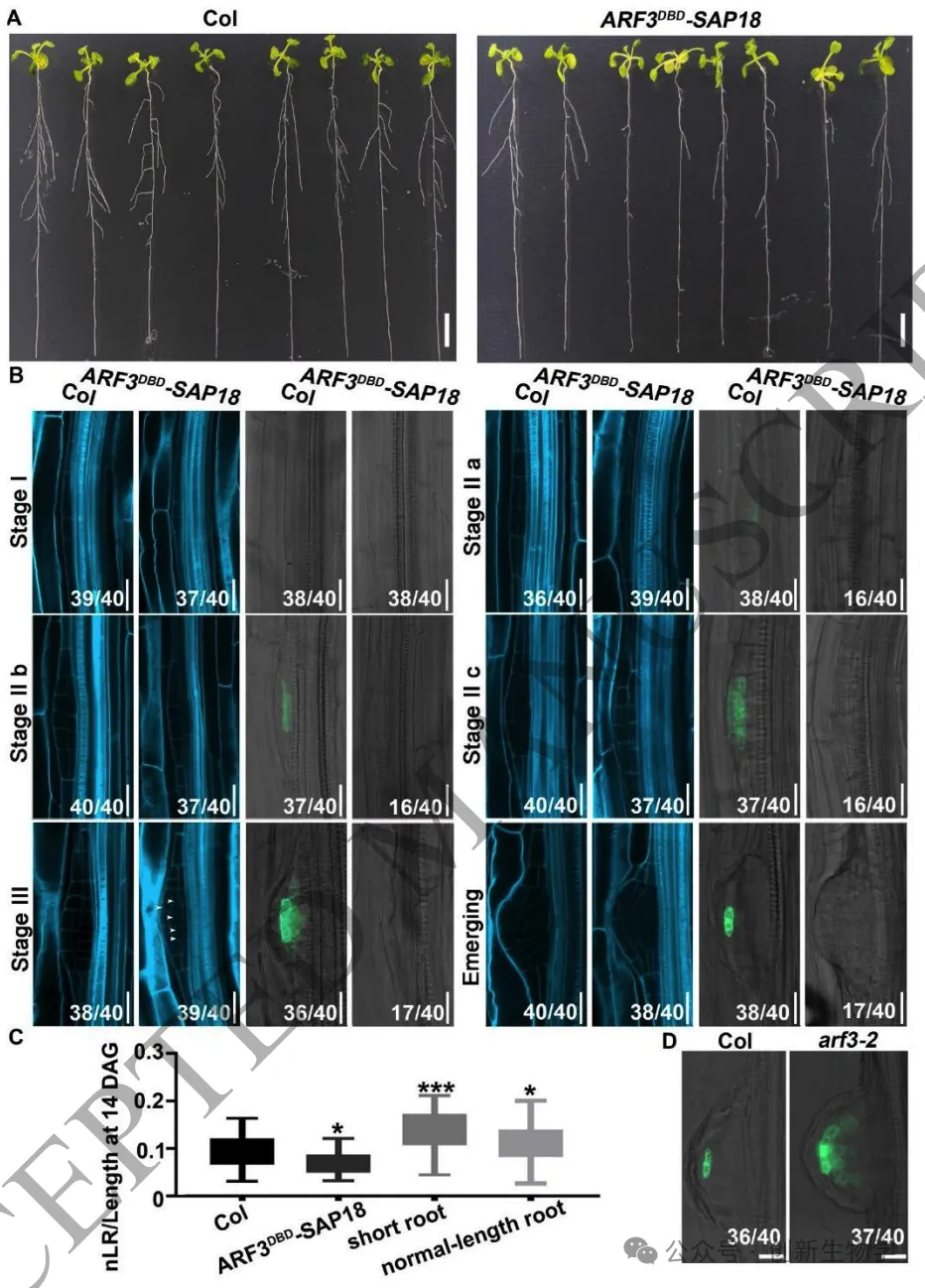
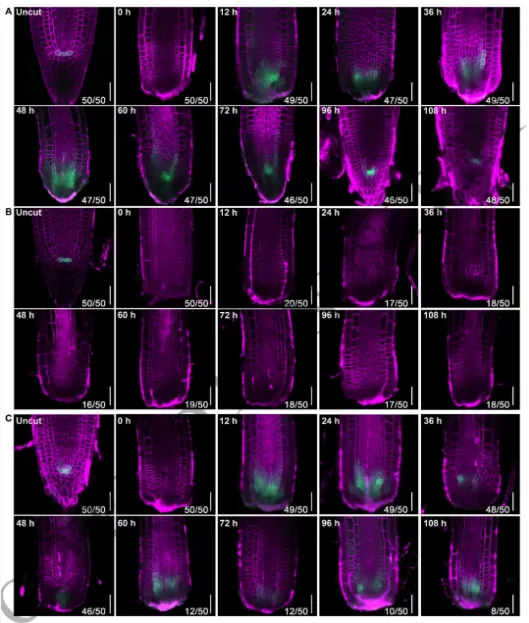
Plant Cell | 山东农业大学张宪省/桑亚林团队揭示根系干细胞龛形成的生长素响应机制
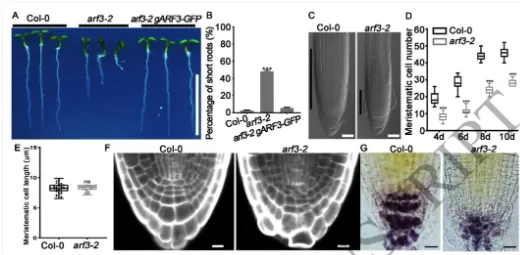
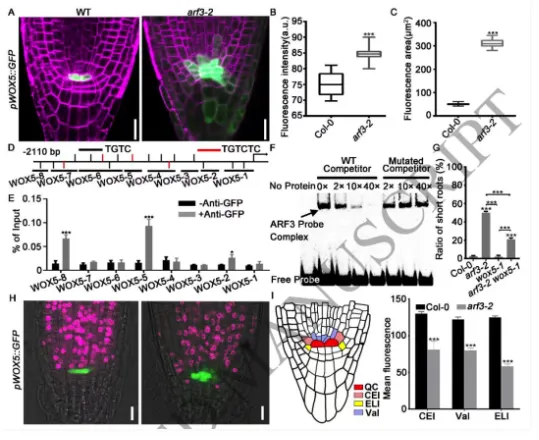
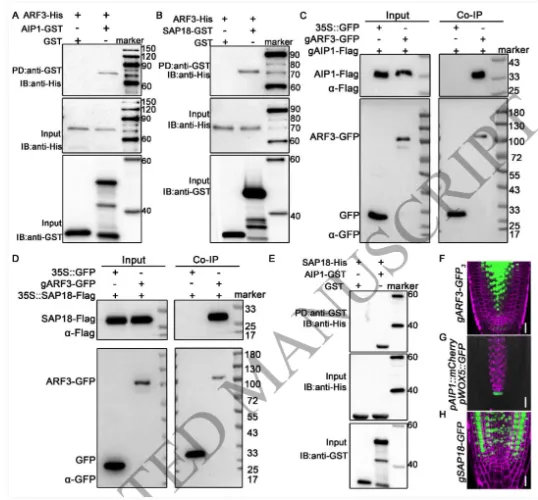
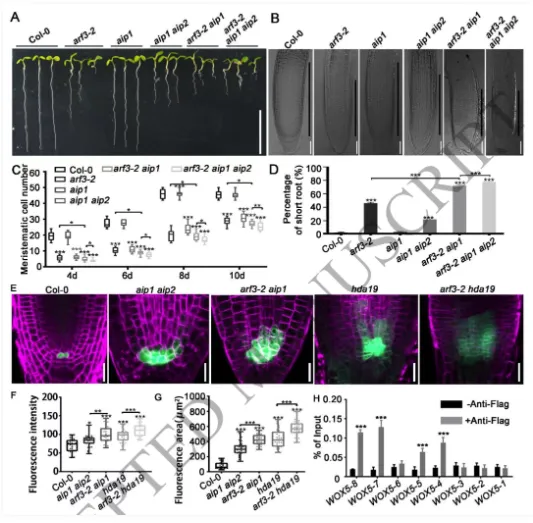
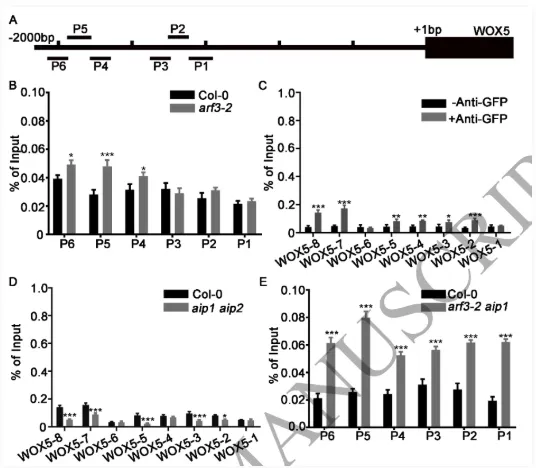
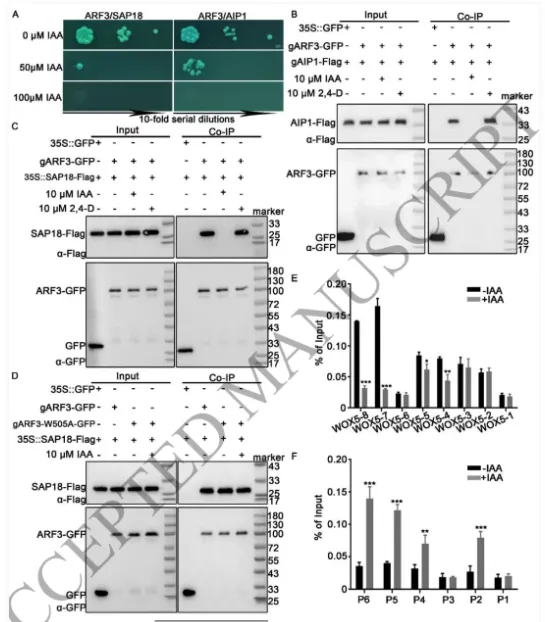
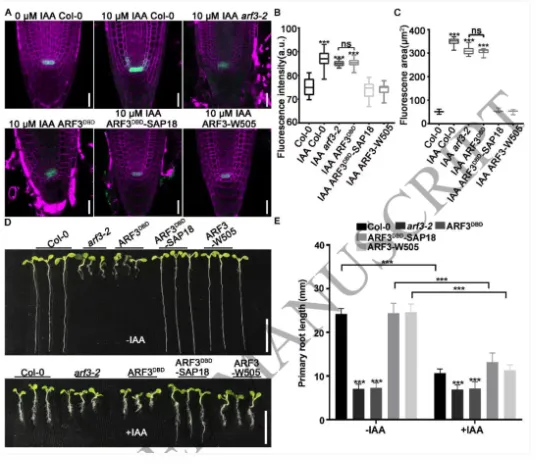
根尖干细胞龛(Stem cell niche, SCN)是植物根系持续生长和器官发生的源泉,由静止中心(Quiescent center, QC)及其相邻的干细胞组成。其中,转录因子WOX5是定义QC身份和维持干细胞不分化状态的“定海神针”,其在 QC 中的特异表达模式对于根系的正常发育至关重要。长期以来,生长素(Auxin)的动态分布梯度是干细胞龛建成与重建的指挥棒,但生长素究竟如何通过特定的分子模块在不同细胞类型中精确开启或关闭WOX5的转录,其底层的生化开关机制仍不完全清楚。研究团队以此为切入点,试图探寻连接生长素信号与 WOX5 空间限制表达之间的“失落环节”。2026年4月10日,山东农业大学生命科学学院、林学院、小麦育种全国重点实验室张宪省教授与桑亚林教授团队在国际植物学顶尖期刊《The Plant Cell》上在线发表(最终校稿还未提交)了题为“The Arabidopsis ARF3-AIP1/2-SAP18 module specifies the root stem cell niche in response to auxin”的研究论文。研究首次鉴定到一个由非典型生长素响应因子ARF3、适配蛋白AIP1/2和共阻遏蛋白SAP18构成的转录抑制复合体,并揭示了其通过响应生长素水平,介导组蛋白去乙酰化修饰,从而在空间上精准限制核心发育基因WOX5的表达范围,以维持根系干细胞龛稳态的分子机制。这一发现不仅阐明了生长素信号如何跨越到表观遗传层面调控干细胞命运,也为通过根系构型改良提升作物养分吸收效率提供了重要的理论支撑。为探究ARF3在根干细胞龛维持中的作用,研究以拟南芥Col-0为野生型,利用arf3-2突变体进行表型分析,发现arf3-2突变体中46%的幼苗表现出短根表型,其根分生组织区长度、皮层细胞数量显著减少,细胞排列异常,静止中心和根冠柱结构紊乱,而ARF3基因回补可恢复该表型,表明ARF3对根分生组织维持和干细胞龛稳态至关重要,且分生组织大小异常主要源于细胞增殖减少而非细胞伸长改变。为明确ARF3调控根生长的分子靶点,研究通过ChIP-qPCR和EMSA实验证实,ARF3可直接结合WOX5基因的启动子区域,负调控WOX5的转录;利用arf3-2与wox5-1突变体杂交获得双突变体,发现其短根表型发生率较arf3-2单突变体降低约50%,表明ARF3通过调控WOX5表达影响根生长;EdU染色实验显示,arf3-2短根突变体近侧干细胞的细胞增殖能力显著下降,而正常长度根中细胞增殖无明显异常,说明ARF3通过限制WOX5在QC中的表达,保障近侧干细胞的增殖能力。为探究ARF3调控WOX5转录的分子机制,研究通过酵母双杂交筛选ARF3相互作用蛋白,鉴定出未知功能的LRR蛋白AIP1及其同源蛋白AIP2,以及SIN3-HDAC复合物组件SAP18;通过一对一酵母双杂交、双分子荧光互补(BiFC)、pull-down和Co-IP实验,证实ARF3可与AIP1/2、SAP18直接相互作用,且AIP1与SAP18之间也存在相互作用;ARF3的中间区域(MR)是其与AIP1/2、SAP18结合的关键结构域;荧光报告株系分析显示,ARF3、AIP1/2与SAP18在近侧干细胞和中柱中共表达,表明三者可在WOX5抑制发生的细胞中形成蛋白复合物。为明确AIP1/2和SAP18在根干细胞龛调控中的作用,研究分析了相关突变体表型,发现aip1和aip2单突变体无明显表型,而aip1 aip2双突变体有16%表现出短根表型,表明AIP1与AIP2功能冗余;arf3-2 aip1双突变体和arf3-2 aip1 aip2三突变体的短根表型较arf3-2单突变体更严重,且WOX5表达的增强和扩张更显著;hda19突变体(HDAC家族成员)有38%表现出短根表型,其WOX5表达模式与arf3-2突变体相似,且hda19 arf3-2双突变体中WOX5表达扩张更明显;ChIP-qPCR实验证实AIP1和SAP18均可结合WOX5启动子,表明AIP1/2和SAP18参与WOX5表达调控和根干细胞龛维持。AIP1/2和SAP18参与调控WOX5表达和SCN维持为验证ARF3-AIP1/2-SAP18模块通过组蛋白乙酰化调控WOX5表达,研究通过ChIP-qPCR实验发现,arf3-2突变体中WOX5启动子区域的组蛋白H3乙酰化水平显著升高,表明ARF3可维持WOX5启动子区正常的组蛋白乙酰化状态;aip1 aip2双突变体中SAP18与WOX5启动子的结合能力显著下降,且ARF3与SAP18的相互作用消失,arf3-2 aip1双突变体中WOX5启动子区组蛋白H3乙酰化水平进一步升高;综上,ARF3通过直接结合WOX5启动子,在AIP1/2的协助下招募SAP18,进而介导组蛋白去乙酰化,协同抑制WOX5转录。ARF3-AIP1/2-SAP18模块不依赖于基于TIR1/AFB的经典生长素信号通路,促进生长素响应的 WOX5 表达。ARF3-AIP1/2-SAP18模块介导组蛋白H3乙酰化为探究该模块对生长素的响应特性,研究发现外源IAA可破坏ARF3与AIP1、SAP18的相互作用,且该作用依赖于ARF3的W505位点;IAA处理可抑制SAP18与WOX5启动子的结合,提高WOX5启动子区组蛋白H3乙酰化水平,导致WOX5异位表达,与arf3-2和aip1 aip2突变体表型一致;内源性生长素升高的突变体中WOX5转录水平显著升高;此外,pARF3::ARF3DBD-SAP18融合蛋白可抵抗IAA的抑制作用,缓解高生长素条件下的根生长抑制,表明高浓度生长素通过破坏ARF3-AIP1/2-SAP18复合物,促进WOX5表达,抑制根伸长。ARF3与AIP1/2-SAP18的相互作用响应生长素水平为进一步验证该模块在生长素响应中的功能,研究分析了不同株系在IAA处理后的表型和WOX5表达,发现IAA处理可导致野生型WOX5表达扩张,而arf3-2突变体、arf3-2 pARF3::ARF3DBD株系、ARF3DBD-SAP18株系和arf3-2 pARF3::ARF3(W505A)株系的WOX5表达对IAA不敏感;ARF3DBD-SAP18株系在IAA处理下的根生长抑制程度显著低于野生型,而仅表达ARF3的DBD结构域无法恢复arf3-2突变体的表型,表明ARF3的C端蛋白相互作用区域对其功能至关重要,该模块以生长素依赖的方式调控WOX5表达和根伸长。ARF3-AIP1/2-SAP18模块响应生长素水平调控WOX5表达和根伸长为探究该模块在根干细胞龛重塑中的作用,研究分析了侧根形成过程,发现ARF3DBD-SAP18株系的侧根密度显著降低,其侧根原基在Ⅱ、Ⅲ期出现异常细胞分裂平面;野生型侧根原基中WOX5在Ⅱ期开始表达,Ⅲ期定位于中心区域,而ARF3DBD-SAP18株系40%的侧根原基在Ⅱ、Ⅲ期无WOX5表达;arf3-2突变体(尤其是短根个体)的侧根密度显著升高,且其侧根原基中WOX5表达横向扩张,表明该模块通过调控WOX5表达参与侧根形成。ARF3-AIP1/2-SAP18模块调控侧根形成在根尖再生实验中,野生型根尖切断后96%可完成再生,而ARF3DBD-SAP18株系、arf3-2短根株系和arf3-2 aip1双突变体的再生失败率分别为36%、16%和22%;野生型再生过程中,WOX5在切断后12 h在切口上方的内皮层和中柱细胞中异位表达,60 h定位于新形成的QC;ARF3DBD-SAP18株系再生失败的根尖中无WOX5表达,而arf3-2突变体中WOX5异位表达持续至96 h仍未定位于QC,表明该模块的解离是WOX5诱导和根尖再生的必要条件,且其可在再生过程中限制WOX5表达。ARF3-AIP1/2-SAP18模块参与根尖再生研究揭示了ARF3-AIP1/2-SAP18模块作为生长素响应开关,调控根干细胞龛维持与重塑的分子机制,填补了抑制型ARF在根干细胞龛调控中的研究空白,研究不仅完善了植物根干细胞龛稳态调控的分子机制,还为理解抑制型ARF的转录调控模式提供了新的视角,同时为作物根发育的遗传改良提供了理论依据和候选基因;此外,AIP1/2在陆生植物中的保守性提示其可能在植物登陆后与ARF协同进化,为研究植物生长素信号通路的进化提供了新线索。论文来源:https://doi.org/10.1093/plcell/koag108