文献分享丨中国农业大学李金杰团队揭示水稻CTB5自然变异促进高原粳稻冷适应性的遗传基础和分子机制
- 2026-06-21 10:31:10

英文题目:Natural variation of CTB5 confers cold adaptation in plateau japonica rice
中文题目:CTB5的自然变异赋予高原粳稻抗寒能力
期刊名称:Nature Communications
影响因子:14.7
作者单位:中国农业大学
DOI:https://dx.doi.org/10.1038/s41467-025-56174-5
研究背景
研究路线
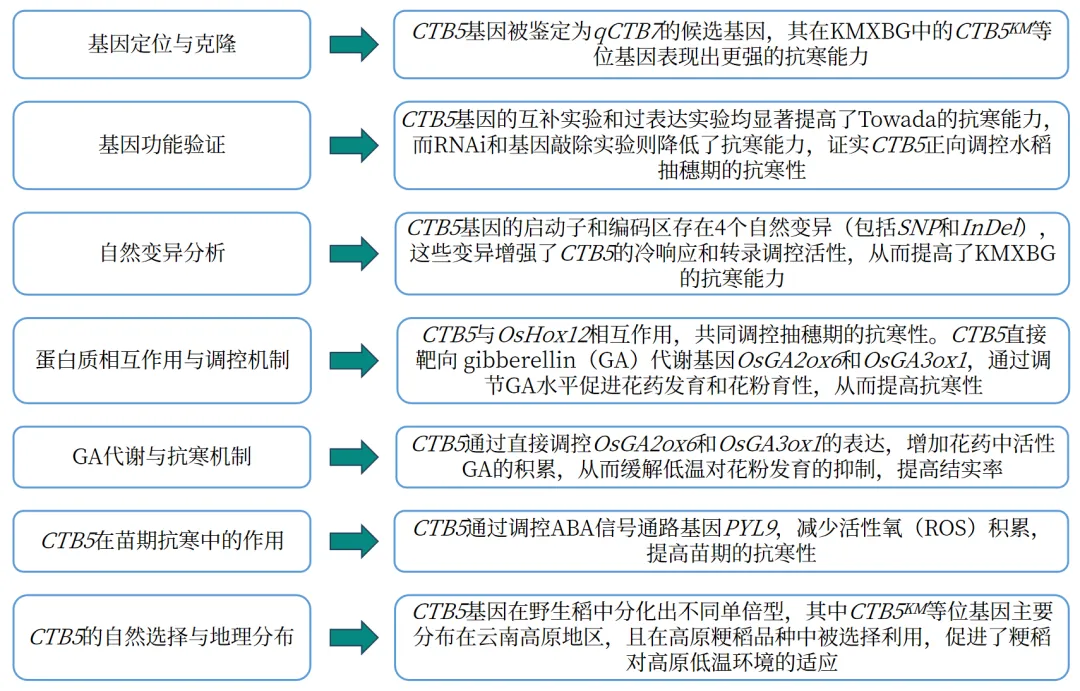
研究结果
01
CTB5在孕穗期积极调节抗寒性
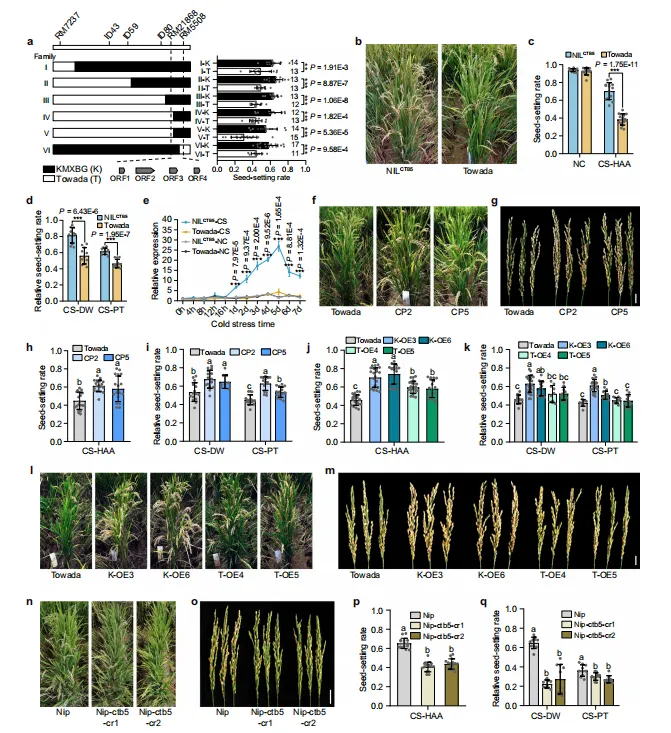
图1.CTB5对孕穗期的抗寒性至关重要。(a)qCTB7的纯合重组体及其抗寒性能。数据代表平均值±标准差。在条形图上标明了用于表型的个体的数量(n)。(b)高海拔地区(CS-HAA)寒冷胁迫下NILCTB5和Towada的田间表现比较。(c、d)正常条件(NC)和CS-HAA条件下(c)以及深水(CS-DW)寒冷胁迫下和植物生长室(CS-PT)条件下(d)NILCTB5和Towada的结实率。数据代表平均值±标准差(NC和CS-HAA为n=15株植物,CS-DW和CS-PT为n=11株植物)。(e)NILCTB5和Towada在寒冷处理期间不同时间点的CTB5相对表达水平。数据代表平均值±SD(n=3个生物学重复)。(f、g)CS-HAA处理下Towada和CTB5互补系的田间表现(f)和穗(g)比较。比例尺g=2cm。(h、i)CS-HAA(n=20/18/18株)(h)CS-DW(n=15/15/10株)和CS-PT(n=15/15/15株)(i)下Towada和CTB5互补系的结实率。(j、k)CS-HAA(n=23/22/19/22/18株)(j)CS-DW(n=13/24/15/10/10株)和CS-PT(n=10/20/12/12/12株)(k)下Towada和CTB5过表达系的结实率。(l、m)CS-HAA胁迫下Towada和CTB5过表达系的田间表现(l)和穗(m)比较。比例尺m=2cm。(n)CS-HAA胁迫下Nipponbare和CTB5敲除系的田间表现(n)和穗(o)比较。比例尺o=2cm。(p、q)CS-HAA(n=15株植物)(p)CS-DW(n=10株植物)和CS-PT(n=10株植物)(q)下Nipponbare和CTB5敲除系的结实率。在(a、c-e)中,星号表示双尾Student’s t检验的统计学意义(**P<0.01,***P<0.001,数据代表平均值±标准差)。在(h–k,p、q)中,不同的小写字母表示通过Duncan多重范围检验的单因素方差分析在P=0.05处具有统计学显著差异,误差线表示平均值±标准差。
02
CTB5通过靶向OsGA2ox6和OsGA3ox1来调节花药中的GAs水平
外源应用试验表明,低温下施用赤霉素(GA)可提高CTB5敲除植物和过表达植物的结实率(图2a、b)。为了研究CTB5是否参与GA途径,我们用HPLC-MS/MS测量了正常和寒冷胁迫条件下CTB5转基因型和野生型植物花药中各种GA成分的水平。GA1、GA3、GA4和GA7是水稻中主要的活性GA,其组织分布不同:GA1和GA3主要在营养器官中,而GA4和GA7主要在生殖器官中。此外还观察到花药中GA4和GA7的含量明显高于GA1和GA3(图2c、d)。在正常条件下,CTB5敲除型、过表达型和野生型植物之间没有发现活性GA水平的显著差异(图2c、d)。然而,在16℃下冷处理7天后,所有植物类型的GA4和GA7水平均显著降低,敲除植物的水平低于Nipponbare,而过表达植物的水平高于Towada(图2c、d)。同样,中间前体GA9和GA15以及产物GA34的水平在CTB5敲除植物中低于Nipponbare,在过表达植物中高于Towada(图2e-h)。相反,GA1、GA3、它们的中间前体GA19和GA20以及产物GA8的水平在两种条件下始终较低,CTB5敲除型、过表达型和野生型植物之间的水平相似(图2c-h)。这些结果表明CTB5介导GA稳态并促进寒冷胁迫下花药中生物活性GA的积累。
为了阐明CTB5在孕穗期调控GA介导的抗寒性的机制,我们使用转录组分析和qRT-PCR鉴定出CTB5通过调控基因OsGA2ox6和OsGA3ox1从而调控GA介导的抗寒性。为了测试CTB5是否直接调节OsGA2ox6和OsGA3ox1,我们分析了这两个基因的启动子并确定了保守位点(TAATTA)基序。酵母单杂交(Y1H)试验表明CTB5与OsGA2ox6和OsGA3ox1的启动子片段直接相互作用(图2i,m)。DNA电泳迁移率变动试验(EMSA)表明CTB5MBP融合蛋白可以与含有TAATTA基序的探针结合,但不能与突变的探针结合(图2j,n)。染色质免疫沉淀(ChIP)分析证实,只有含有TAATTA基序的片段显著富集(图2k,o)。双荧光素酶报告基因检测显示,CTB5抑制OsGA2ox6并激活OsGA3ox1,且CTB5KM的调控作用强于CTB5Tow(图2l,p)。此外,OsHox12和CTB5共表达显著抑制了OsGA2ox6的LUC表达并增强了OsGA3ox1的LUC表达,表明OsHox12促进了CTB5对这两个基因的调控作用(图2q-t)。
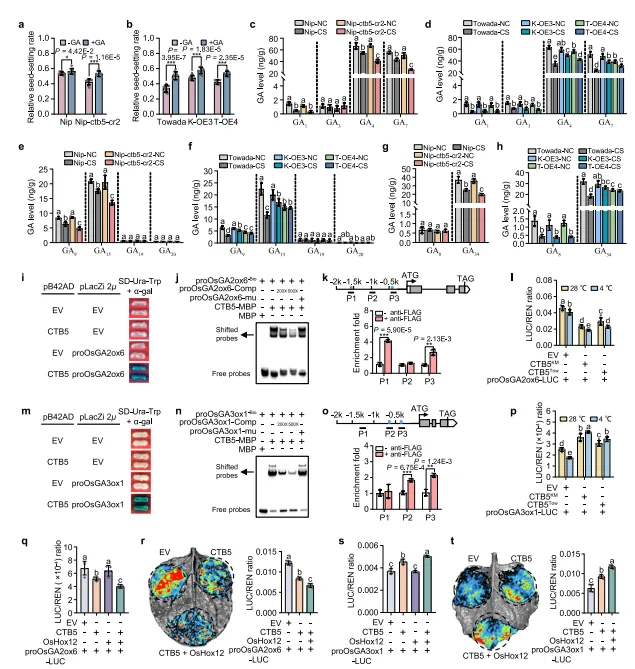
图2.CTB5通过直接调节OsGA2ox6和OsGA3ox1来调节花药中的赤霉素(GA)水平。(a、b)在施用和不施用GA的情况下暴露于冷胁迫的CTB5敲除(a)和过表达(b)植物的结实率。数据代表平均值±标准差(a中n=11/9、10/9株植物,b中n=10/10、10/10、7/7株植物)。(c-h)在正常条件(NC)和冷胁迫(CS)条件(16℃,7天)下CTB5敲除和过表达植物正在发育的花药中的GA含量。数据代表平均值±标准差(n=3个生物学重复)。(i、m)酵母单杂交试验测试CTB5与OsGA2ox6(i)和OsGA3ox1(m)启动子的结合。EV,空载体。(j、n)EMSA检测表明CTB5与OsGA2ox6(j)和OsGA3ox1(n)启动子的TAATTA基序结合。(k、o)ChIP-qPCR检测表明CTB5在体内与OsGA2ox6(k)和OsGA3ox1(o)启动子结合。数据代表平均值±标准差(n=3个生物学重复)。(l、p)双荧光素酶报告基因检测证实CTB5与水稻原生质体中的OsGA2ox6(l)和OsGA3ox1(p)启动子结合。EV,空pGreenII62-SK载体。数据代表平均值±标准差(l为n=5个生物学重复,p为n=4个生物学重复)。(q-t)双荧光素酶报告基因检测显示,OsHox12增强了CTB5对水稻原生质体(q,s)和烟草叶片(r,t)中OsGA2ox6和OsGA3ox1的调控作用。EV,空pGreenII62-SK载体。数据代表平均值±标准差(q和s的n=4个生物学重复,r和t的n=5个生物学重复)。在(i–t)中,显示了来自至少两个独立实验的代表性实验。在(a、b、k、o)中,星号表示通过双尾Student’s t检验的统计学显着性(*P<0.05,**P<0.01,***P<0.001)。在(c–h,l,p–t)中,不同的小写字母表示通过单因素方差分析和Duncan多范围检验在P=0.05处具有统计学显着差异。
03
CTB5调控冷胁迫下绒毡层发育和花粉育性
为了阐明CTB5调控孕穗期抗寒性的组织学机制,我们检测了参与绒毡层发育的关键基因的表达,包括EAT1、LAP6和DPW3。在低温胁迫下,这些基因的表达在Nip-ctb5-cr2中显著低于Nipponbare,在K-OE3中显著高于Towada。然而,在正常条件下,Nip-ctb5-cr2和Nipponbare之间,或K-OE3和Towada之间这些基因的表达水平无显著差异(图3a)
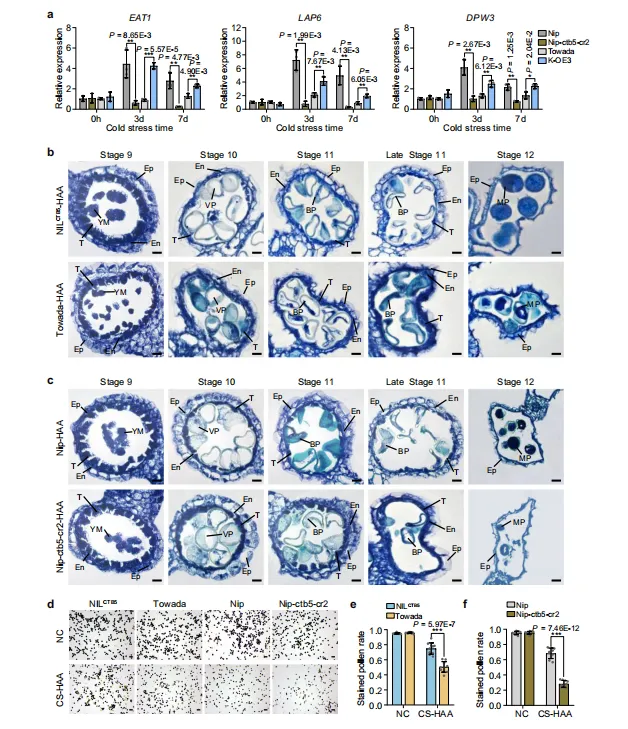
图3.CTB5在寒冷胁迫下调节绒毡层发育和花粉育性。(a)在正常和寒冷胁迫条件下,CTB5敲除和过表达系中EAT1、LAP6和DPW3的相对表达水平。数据代表平均值±标准差(n=3个生物学重复)。(b、c)在高海拔地区寒冷胁迫(CS-HAA)条件下,NILCTB5和Towada(b)以及Nipponbare和CTB5敲除植物(c)花药发育的组织学分析。比例尺=10μm。Ep,表皮细胞层;En,内皮细胞层;YM,小孢子;VP,液泡花粉;BP,双细胞花粉;MP,成熟花粉;T,绒毡层。(d)在正常条件(NC)和CS-HAA下对NILCTB5、Towada、Nipponbare和CTB5敲除植物的花粉进行染色。比例尺=100μm。(e、f)NC和CS-HAA条件下NILCTB5和Towada(e)以及Nipponbare和CTB5敲除植物(f)的染色花粉率。数据代表平均值±标准差(n=10个生物学重复)。在(a、e、f)中,星号表示双尾Student’s t检验的统计显著性(*P<0.05,**P<0.01,***P<0.001)。
讨论与总结
Guo, H., Gao, S., Li, H. et al. Natural variation of CTB5 confers cold adaptation in plateau japonica rice. Nature Communications 16, 1032 (2025). https://doi.org/10.1038/s41467-025-56174-5.

END
推 荐 阅 读
2、文献分享丨山东农业大学李媛媛团队解析氮素对苹果重要外在品质—表皮蜡质的调控机制
3、合作文章丨中科院武汉植物园杨美团队揭示莲子长寿的内在分子机制

随机文章
-
 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
- IF 55.8:合成生物学颠覆农业!植物 + 微生物工程解锁未来农业与环境新范式
- 260413青旗农业 脱毒种苗 助您增收
- 溯源农耕文明,探秘农业科技|西农博览园“农脉传承·科创赋能·科普铸魂”主题系列研学活动(二)
- 华中农业大学招聘公告!报名中!
- 项目采购|北湖区石盖塘街道江口村农业基础设施建设项目采购公告
- 农艺师证书权威解析!农业农村部直属发证,官网可查,月薪 8K+,新农人必备!
- 湖北国家级智慧农业示范基地项目盛大启航
- 订单农业:让农业从靠天吃饭到靠订单赚钱
- 以工业化思维做大做强现代农业
- 赛乌素嘎查新农人:敢闯敢试 让智慧农业结出“甜蜜果” | 走一线 改文风
