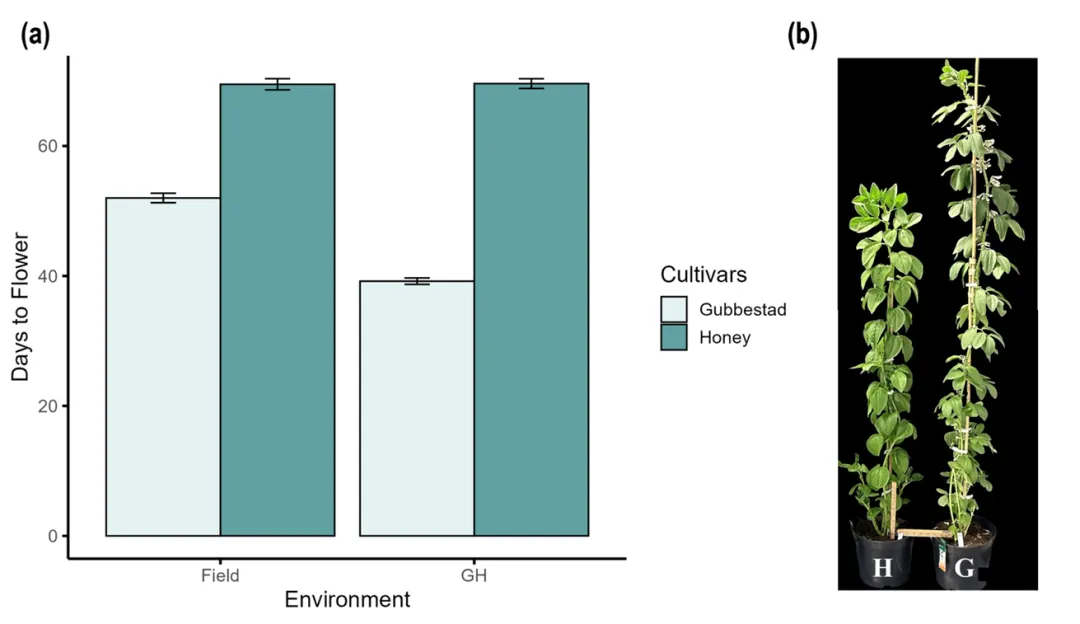
研究人员选取早花品种Gubbestad和晚花品种Honey为试验材料,人工气候箱培养结果显示,Gubbestad在播种后39天(DAS)开花,Honey开花时间则推迟至68DAS,二者在田间和温室环境下均表现出稳定的开花时间差异(图 1)
图1 室外与温室环境中不同植物材料首次开花出现天数比对
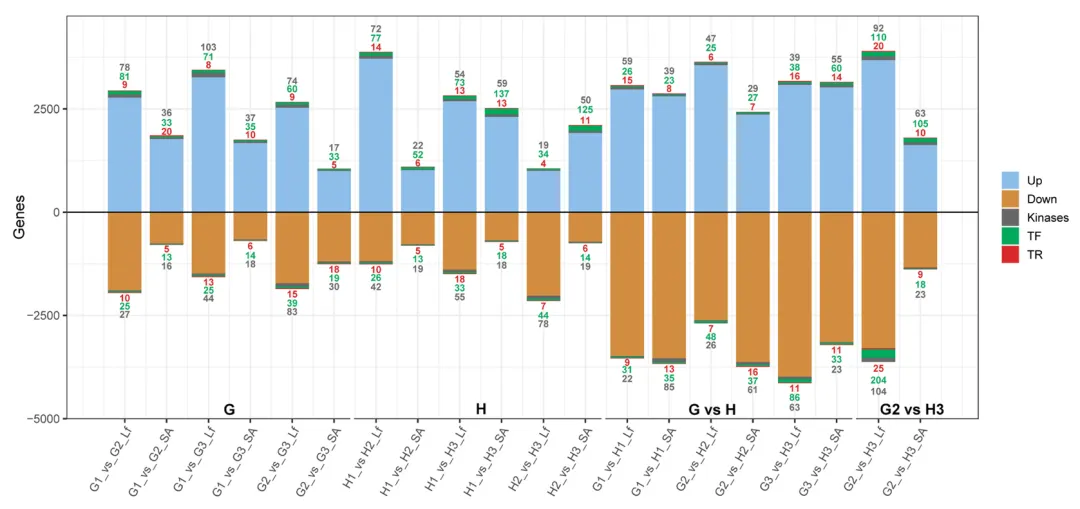
转录组测序共获得平均每样品 7276 万条高质量读段,75% 的读段可唯一比对到蚕豆参考基因组,数据深度满足后续分析要求。以 | log₂(倍变化)|≥1、假发现率(FDR)<0.05 为标准,全样本共鉴定出 16864 个差异表达基因(DEGs),不同品种、发育阶段、组织间的 DEGs 数量和调控类型存在显著差异(图 2)。
图2 不同发育阶段、组织及栽培品种间的差异表达基因
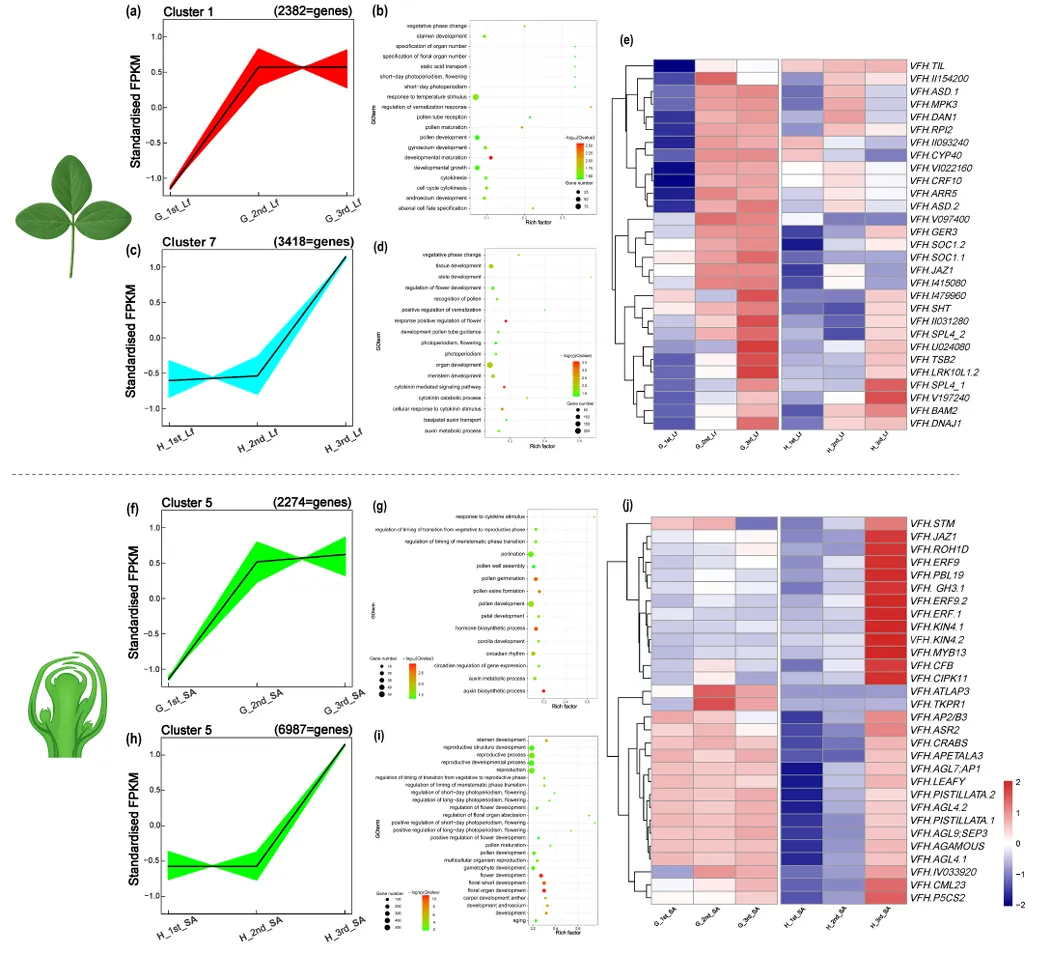
对 DEGs 进行 GO 富集分析,结果显示蚕豆开花诱导的基因功能富集存在品种特异性时序和组织特异性分工:Gubbestad 在 39 DAS 时,叶片已富集赤霉素(GA)合成 / 代谢、光信号响应相关通路,茎尖富集花分生组织生长、营养 - 生殖阶段转换通路;而 Honey 上述开花相关通路均延迟至 68 DAS 才显著富集,且茎尖的富集程度更高。对 DEGs 进行 K-means 聚类分析(最优聚类数为 12),结果显示早花、晚花品种的开花相关基因聚为特异性表达簇,Gubbestad 的开花关键基因在 39 DAS 即出现表达峰值,而 Honey 的同源基因表达峰值推迟至 68 DAS;叶片和茎尖的开花相关基因聚类无明显重叠,表明叶片主要介导环境信号感知,茎尖主导成花转换与花发育的执行(图 3)。
图3 K均值聚类分析与差异表达基因的功能富集分析
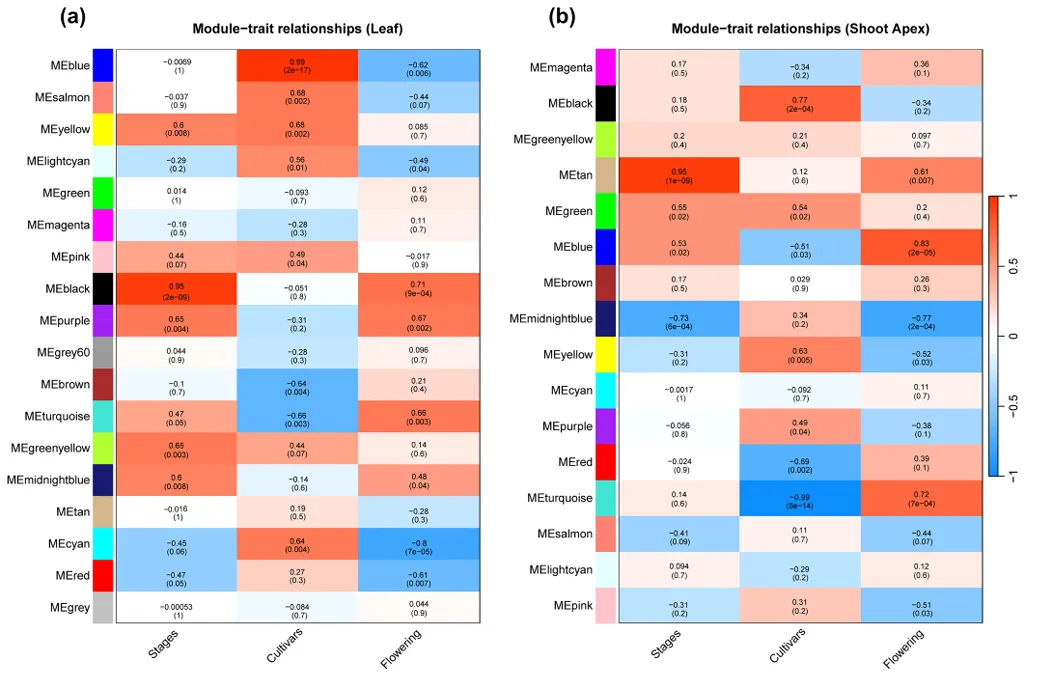
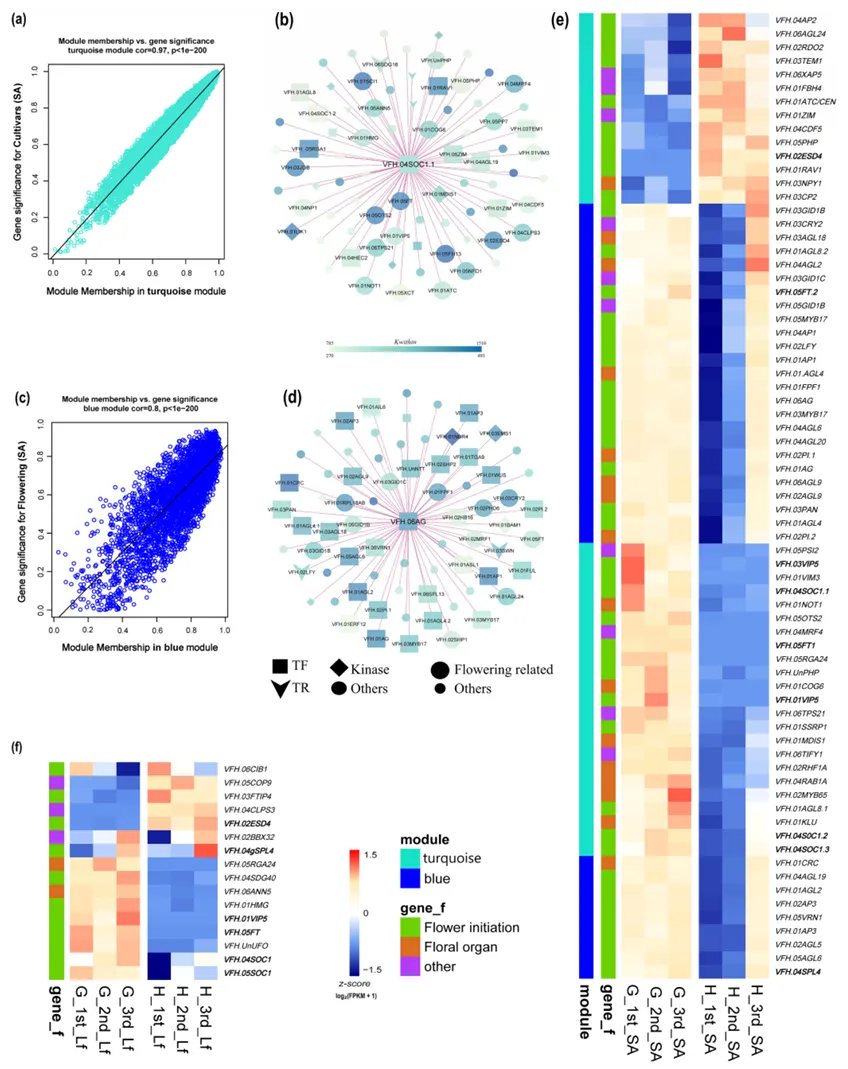
以 FPKM>1 的基因为基础,构建叶片和茎尖的加权基因共表达网络(WGCNA),确定最优软阈值为 12,最终在叶片中鉴定出 18 个共表达模块,茎尖中鉴定出 16 个共表达模块,模块与开花时间、品种、发育阶段的关联存在显著组织特异性(图 4)。
图4 叶片(a)与SA(b)中的模块特性关系
叶片中MEblack 模块与开花诱导高度相关,富集昼夜节律、光刺激响应、温度响应等环境信号感知相关功能;
茎尖中MEturquoise 模块(与品种高度相关,r²=-0.99)和MEblue 模块(与开花时间高度相关,r²=0.83)为开花调控核心模块,其中 MEturquoise 模块的核心枢纽基因为VFH.04SOC1.1,MEblue 模块的核心枢纽基因为VFH.06AG(图 5);
枢纽基因的共表达网络分析显示,SOC1 与 FT、VIP5、CDF5、TEM1 及 GA 信号通路基因紧密关联,AG 与 AP1、AP3、PI、GID1B/C、SPL4、CRY2 等花发育和信号通路基因共表达,二者共同构成蚕豆开花诱导的核心调控网络(图 5)。
图5 通过 WGCNA 鉴定的开花相关模块共表达网络分析
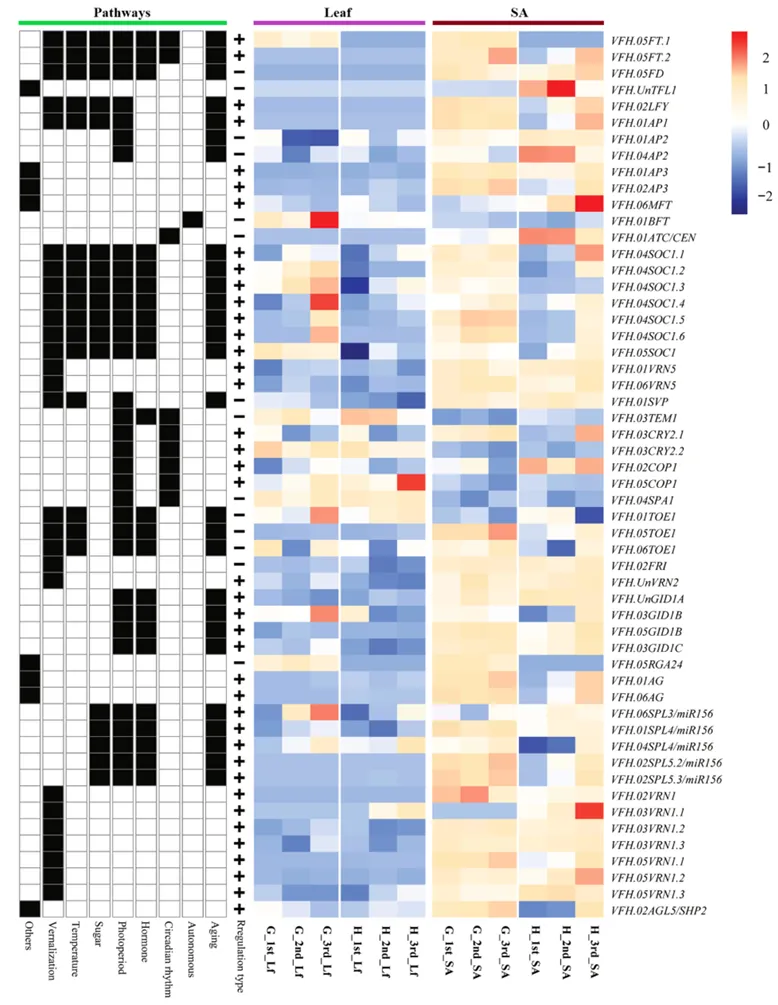
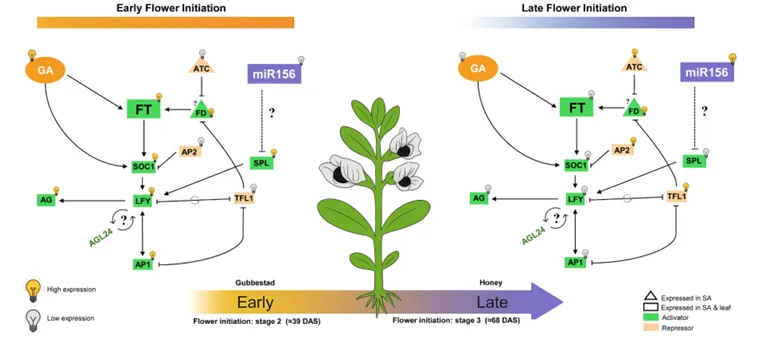
对光周期、GA 信号、年龄途径等核心开花通路基因的表达模式分析,明确了早花、晚花品种的表达差异核心(图 6)。结果显示,FT 同源基因:Gubbestad 的叶片和茎尖中均有 2 个 FT 基因表达,且营养期即启动表达;Honey 仅在茎尖生殖期检测到 1 个 FT 基因表达,且表达量显著低于 Gubbestad;开花抑制基因:Honey 的茎尖在全发育阶段均高表达 TFL1、AP2、ATC、TOE1,而 Gubbestad 中这些基因的表达水平显著下调;开花激活基因:Gubbestad 在 21 DAS 营养期即高表达 SOC1 家族基因、LFY、AP1、SPL4/5 及 GA 受体基因 GID1B,提前启动成花转换;GA 信号通路基因:Gubbestad 的 GA 合成、信号转导相关基因在 39 DAS 已显著上调,而 Honey 的同源基因延迟至 68 DAS 才激活。
图6 拟南芥中参与调控蚕豆叶片及SA开花过程的部分关键基因的表达热图