3.1. QQS表达增强紫花苜蓿的根系分枝与抗旱性
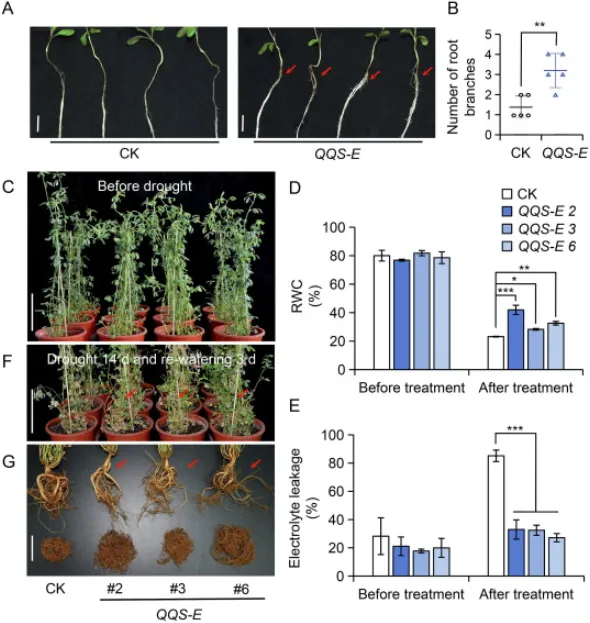
为探究QQS基因的功能,对QQS-E T1代转基因幼苗进行了表型分析和抗旱性测试。结果显示,与对照(CK)相比,表达QQS的植株表现出显著增强的根系分枝能力。经过14天的干旱胁迫处理后,QQS-E植株的萎蔫程度远低于CK植株,其叶片具有更高的相对含水量(RWC)和更低的电解质渗漏率,表现出更强的抗旱性。复水后,QQS-E植株能快速恢复生长,而CK植株恢复有限。这表明QQS的表达能够通过促进根系分枝,进而提高紫花苜蓿的抗旱能力(Fig. 1)。
图1. QQS增强紫花苜蓿的根系分枝和抗旱性。(A, B) 两周龄CK(非转基因)和QQS-E(转基因)幼苗的根系分枝表型及其根分枝数统计分析(n=5)。实验使用QQS-E株系2。红色箭头指示根分枝位置。标尺=1 cm。,P < 0.01(学生t检验)。(C, F) 3月龄CK和QQS-E幼苗在干旱胁迫前以及经历14天干旱胁迫并复水3天后的表型。红色箭头指示恢复良好的分枝位置。标尺=10 cm。(D, E) 干旱胁迫下测试植株叶片的相对含水量(RWC)和电解质渗漏率(n=3)。*,P < 0.05;,P < 0.01;***,P < 0.001(单因素方差分析)。误差棒代表平均值±标准差(SD)。(G) 复水后QQS-E和CK植株的根系表型。红色箭头指示根分枝位置。标尺=5 cm。
3.2. QQS与核定位转录因子MsNF-YC4互作
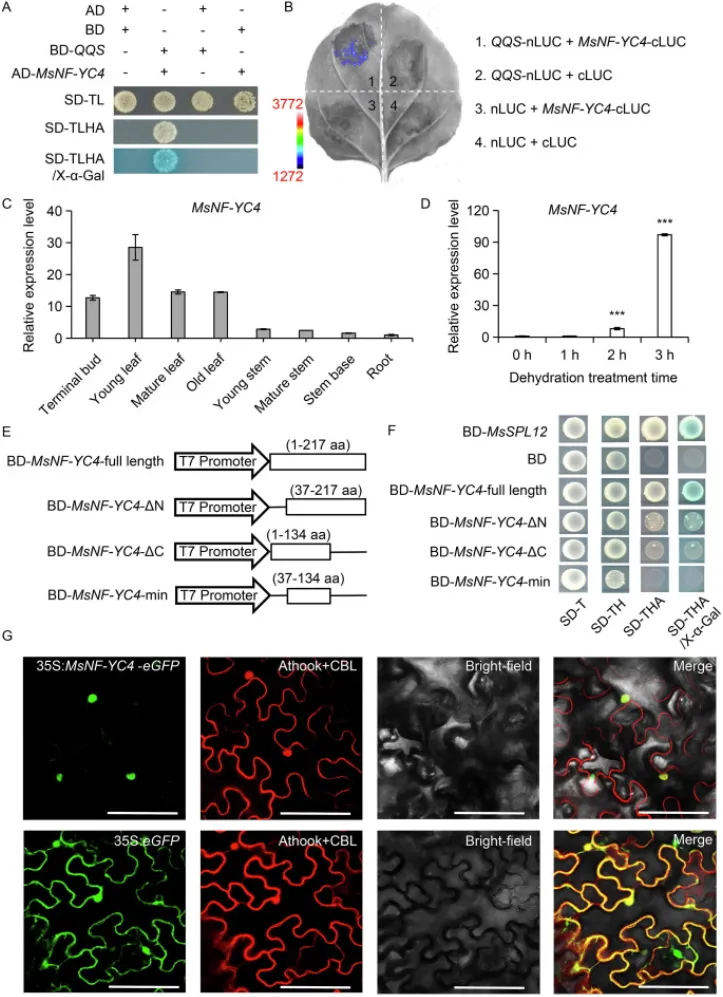
通过酵母双杂交(Y2H)文库筛选,鉴定出MsNF-YC4是QQS在紫花苜蓿中的一个潜在内源互作蛋白,该互作关系通过一对一的Y2H和荧光素酶互补成像(LCI)实验得到进一步证实。RT-qPCR分析表明,MsNF-YC4基因在紫花苜蓿的各个器官中均有表达,且在根部受到脱水处理的显著诱导。亚细胞定位实验显示MsNF-YC4蛋白定位于细胞核。此外,酵母系统中的转录激活分析证实,全长的MsNF-YC4蛋白具有转录激活活性,而其截短体则不具备此功能。这些结果表明,MsNF-YC4是一个响应干旱胁迫的核定位转录因子,并能与QQS蛋白直接互作(Fig. 2)。
图2. QQS与核定位转录因子MsNF-YC4互作。(A) 酵母双杂交(Y2H)分析显示QQS与MsNF-YC4之间的相互作用。SD-TL:SD/-Trp-Leu培养基;SD-THLA:SD/-Trp-Leu-His-Ade培养基;SD-THLA/X-α-Gal:添加了X-α-Gal的SD-THLA培养基。(B) QQS与MsNF-YC4的体内相互作用。(C) MsNF-YC4在紫花苜蓿不同组织中的表达模式。(D) 脱水处理下紫花苜蓿根部MsNF-YC4的表达模式。***,P < 0.001(单因素方差分析)。数值为平均值±标准差。(E, F) 全长(BD-MsNF-YC4-full length)及截短体(BD-MsNF-YC4-ΔN, BD-MsNF-YC4-ΔC, 和 BD-MsNF-YC4-min)MsNF-YC4编码序列在酵母系统中的转录激活分析。SD-T:SD/−Trp;SD-TH:SD/−Trp-His;SD-THA:SD/−Trp-His-Ade;SD-THA/X-α-Gal:添加了X-α-Gal的SD-THA培养基。(G) MsNF-YC4在烟草叶细胞中的亚细胞定位。Athook:核定位标记蛋白;CBL:质膜定位标记蛋白。标尺=100 µm。
3.3. MsNF-YC4增强根系分枝与抗旱性
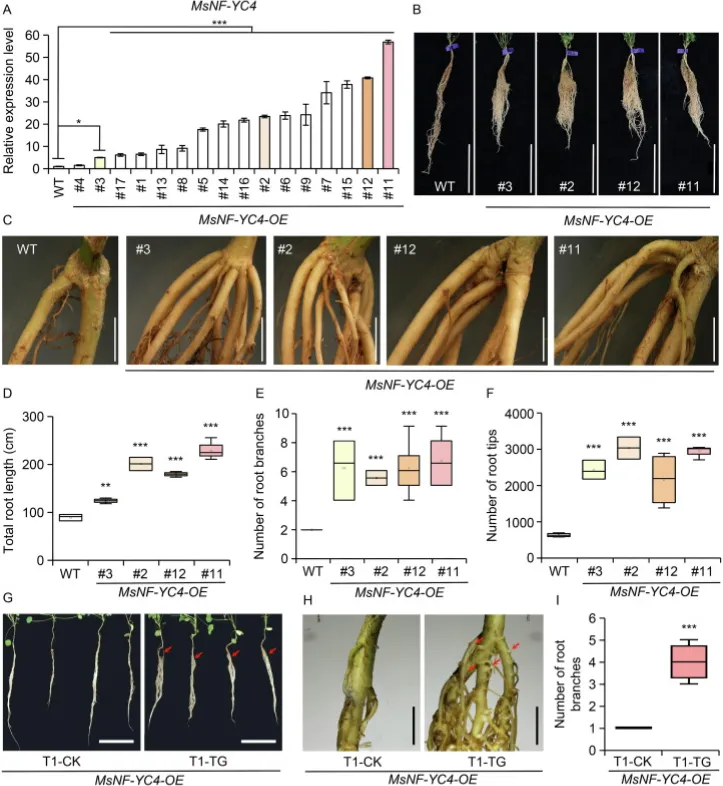
为验证MsNF-YC4在根系发育和抗旱性中的作用,构建了过表达该基因的转基因紫花苜蓿株系(MsNF-YC4-OE)。表型分析显示,与野生型(WT)相比,MsNF-YC4-OE植株具有更庞大的根系、更长的总根长以及显著更多的根分枝和根尖数目,且该多分枝根系的表型在其T1代转基因后代中能够稳定遗传(Fig. 3)。在干旱胁迫下,MsNF-YC4-OE植株表现出更高的存活率、更高的相对含水量和更低的电解质渗漏率。此外,其离体叶片的失水速率更慢,且在PEG模拟干旱处理下气孔开度更小。这些结果有力地证明,过表达MsNF-YC4能够显著促进紫花苜蓿的根系分枝,并提高其抗旱能力(Fig. 4)。
图3. MsNF-YC4调控紫花苜蓿的根系发育。(A) MsNF-YC4-OE植株中MsNF-YC4的相对转录水平(n=3)。,P < 0.05;,P < 0.001(单因素方差分析)。(B, C) 由扦插繁殖的3月龄MsNF-YC4-OE和WT植株的根系结构。标尺分别为10 cm (B) 和 5 mm (C)。(D) MsNF-YC4-OE和WT植株的总根长(n=5)。,P < 0.01;,P < 0.001(学生t检验)。数值为平均值±标准差。(E) MsNF-YC4-OE和WT植株根分枝数的统计分析(n=6)。,P < 0.001(单因素方差分析)。数值为平均值±标准差。(F) MsNF-YC4-OE和WT植株的总根尖数。星号表示经单因素方差分析确定的显著性差异(, P < 0.001)。数值为平均值±标准差。(G, H) 1月龄MsNF-YC4-OE* T1代转基因幼苗和CK植株的根系表型。红色箭头指示根分枝位置。标尺分别为5 cm (G) 和 5 mm (H)。(I) MsNF-YC4-OE植株T1代幼苗的根分枝数。T1-CK:经PCR鉴定的非转基因幼苗;T1-TG:经PCR鉴定的转基因幼苗(n=5)。*,P < 0.001(学生t检验)。数值为平均值±标准差。
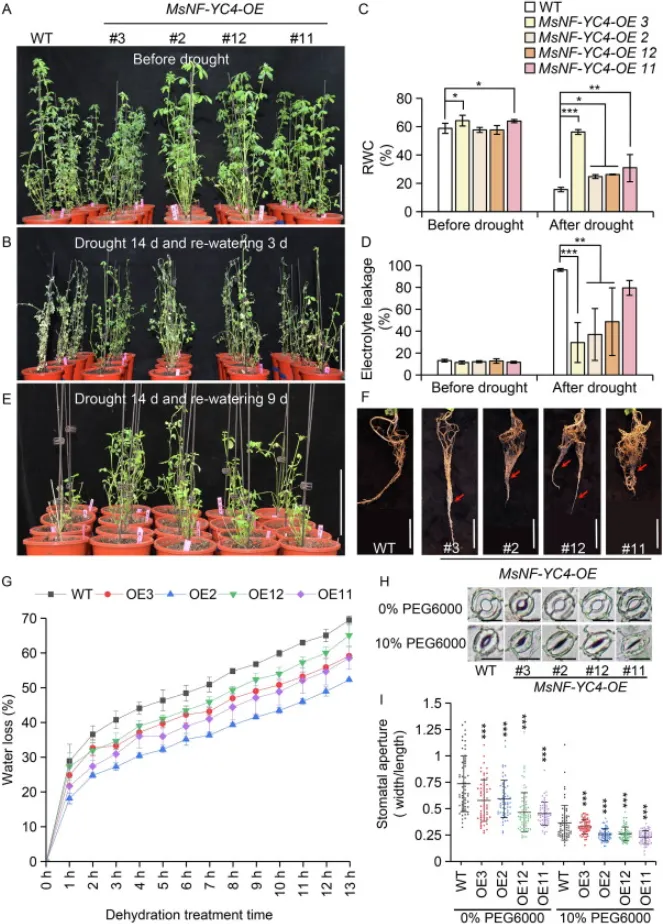
图4. 过表达MsNF-YC4增强紫花苜蓿的抗旱性。(A, B) MsNF-YC4-OE和WT植株在干旱胁迫前及胁迫处理后复水3天的表型。标尺=10 cm。(C, D) 干旱胁迫前后MsNF-YC4-OE和WT紫花苜蓿叶片的相对含水量(RWC)和电解质渗漏率(n=4)。,P < 0.05;,P < 0.01;,P < 0.001(单因素方差分析)。数值为平均值±标准差。(E) 干旱胁迫后复水9天,MsNF-YC4-OE和WT植株新长出的分枝。标尺=10 cm。(F) 复水9天后MsNF-YC4-OE和WT植株的根系表型。红色箭头指示新发育的根。标尺=5 cm。(G) WT和MsNF-YC4-OE植株离体叶片随时间变化的失水率(n=3)。,P < 0.05;**,P < 0.01(单因素方差分析)。数值为平均值±标准差。(H) MsNF-YC4-OE和WT植株在0%或15% PEG6000中暴露1小时后的气孔表型。标尺=10 µm。(I) WT和MsNF-YC4-OE植株在0%或15% PEG6000中处理1小时后的气孔开度(n=60)。,P < 0.05;***,P < 0.001(单因素方差分析)。数值为平均值±标准差。
3.4. 过表达MsNF-YC4调控脱落酸和生长素信号通路
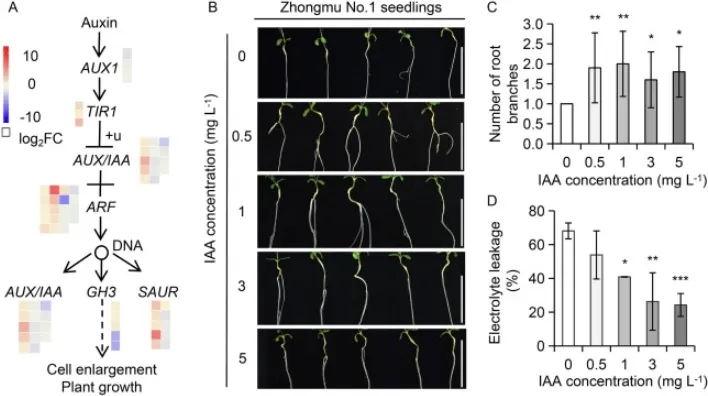
为揭示MsNF-YC4调控根系分枝的分子机制,对MsNF-YC4-OE和WT植株的根部进行了转录组测序(RNA-Seq)。结果共鉴定出21,437个差异表达基因(DEGs)。基因本体论(GO)富集分析显示,这些DEGs显著富集在与胁迫响应和发育相关的生物学过程中,特别是脱落酸信号、生长素代谢、过氧化物清除和根形态建成等通路,其中包括54个与生长素信号通路相关的基因(Fig. 5A)。外源施加生长素(IAA)能够显著促进野生型紫花苜蓿幼苗的根系分枝,并提高其在PEG诱导的干旱胁迫下的耐受性,表现为更低的电解质渗漏率。这表明MsNF-YC4可能通过调控生长素信号通路来影响根系发育和抗旱性(Fig. 5B-D)。
图5. MsNF-YC4-OE根系的转录组分析。(A) 分配至生长素信号转导通路的差异表达基因的热图。颜色强度代表log2FC(倍数变化对数值)。(B, C) 经外源IAA处理的幼苗的植株表型和根分枝数。,P < 0.05;**,P < 0.01(单因素方差分析)。数值为平均值±标准差(n=10)。标尺=3 cm。(D) 在五种浓度的IAA中培养9天后,再用10% PEG6000处理6天的幼苗的电解质渗漏率。星号表示经单因素方差分析确定的显著性差异(, P < 0.05; ***, P < 0.001)。数值为平均值±标准差(n=10)。
3.5. MsNF-YC4激活MsARF8基因的表达
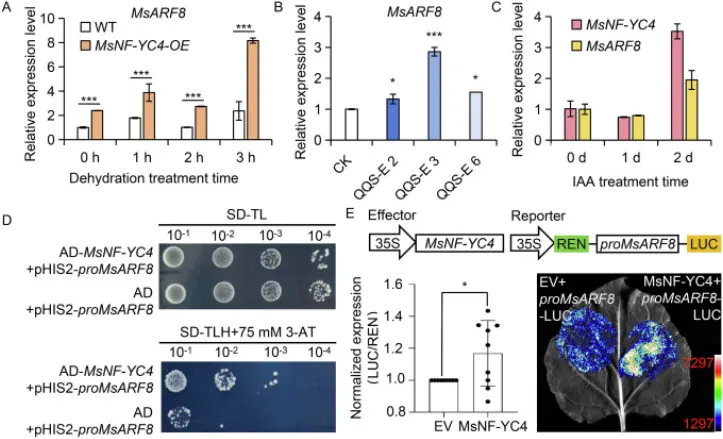
为寻找MsNF-YC4下游与生长素信号相关的靶基因,通过对差异表达基因启动子区域的分析,发现MsARF8、MsARF19、MsABP19b和MsAUX6B等基因的启动子含有NF-Y结合位点。其中,MsARF8的表达在MsNF-YC4-OE和QQS-E的根中均上调,并且其表达能被脱水处理和外源IAA诱导,其响应模式与MsNF-YC4相似(Fig. 6A-C)。酵母单杂交(Y1H)和双荧光素酶(Dual-LUC)报告实验进一步证实,MsNF-YC4蛋白能够直接结合到MsARF8基因的启动子区域,并激活其转录。这些证据表明,MsARF8是MsNF-YC4的一个直接下游靶基因(Fig. 6D, E)。
图6. MsNF-YC4上调MsARF8的转录。(A) 脱水处理下WT和MsNF-YC4-OE根中MsARF8的相对表达水平(n=3)。数值为平均值±标准差。,P < 0.001(单因素方差分析)。(B) CK和QQS-E根中MsARF8的相对表达水平(n=3)。数值为平均值±标准差(n=3)。,P < 0.05;,P < 0.001(单因素方差分析)。(C) 经1 mg L⁻¹ IAA处理的紫花苜蓿幼苗根颈部MsNF-YC4和MsARF8的表达模式。数值为平均值±标准差。(D) 酵母单杂交(Y1H)分析显示MsNF-YC4与MsARF8启动子的结合。SD-TL:SD/-Trp-Leu培养基;SD-TLH + 75 mmol L⁻¹ 3-AT:含75 mmol L⁻¹ 3-AT的SD-TLH培养基。图像于30°C孵育3天后拍摄。(E) 在共转化了MsNF-YC4效应子构建的烟草叶片中,对proMsARF8-LUC报告基因的瞬时激活分析(n=10)。LUC:荧光素酶。,P < 0.05(学生t检验)。
3.6. MsARF8正向调控生长素诱导的根系分枝和抗旱性
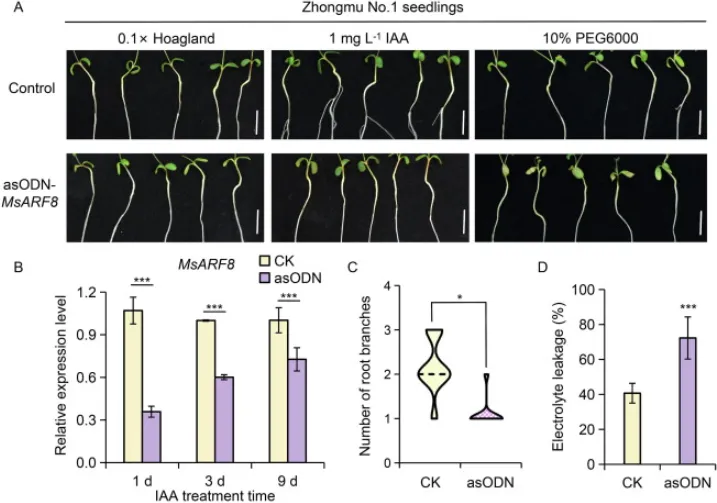
为探究MsARF8的功能,采用瞬时反义寡核苷酸(asODN)技术抑制了其在紫花苜蓿幼苗中的表达。结果显示,抑制MsARF8不仅显著降低了其自身表达水平,还消除了外源IAA对根系分枝的促进作用。在PEG模拟干旱胁迫下,抑制MsARF8的幼苗子叶褐化更为严重,电解质渗漏率显著高于对照组。这表明,MsARF8是调控紫花苜蓿根系分枝和抗旱性的一个关键正向调控因子,介导了生长素诱导的根系发育过程(Fig. 7)。
图7. MsARF8正向调控生长素诱导的根系分枝和紫花苜蓿的抗旱性。(A) 对照组和MsARF8沉默(asODN-MsARF8)幼苗在外源IAA或PEG处理下的表型。标尺=1 cm。(B) 紫花苜蓿幼苗在20 μmol L⁻¹ asODN-MsARF8溶液中浸泡后根颈部MsARF8的相对表达水平(n=3)。,P < 0.001(学生t检验)。数值为平均值±标准差。(C) 外源IAA处理下CK和asODN-MsARF8幼苗的根分枝数(n=10)。,P < 0.05(学生t检验)。(D) 经10% PEG6000处理9天后CK和asODN-MsARF8幼苗的电解质渗漏率(n=5)。*,P < 0.001(学生t检验)。数值为平均值±标准差。
研究的创新性
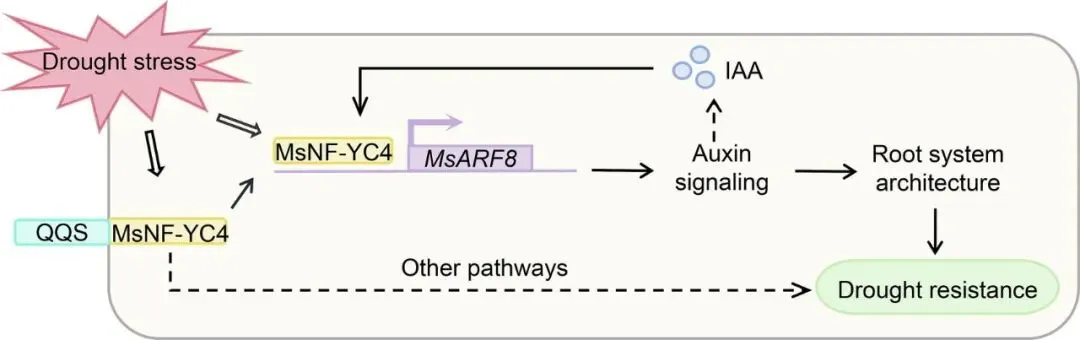
本研究的核心创新之处在于,系统性地阐明了一条在重要牧草紫花苜蓿中,由外源基因QQS介导,通过内源互作蛋白MsNF-YC4及其下游靶基因MsARF8,最终调控根系发育和抗旱性的完整分子通路(QQS → MsNF-YC4 → MsARF8 → 生长素信号 → 根系分枝与抗旱性)。这不仅揭示了QQS在紫花苜蓿中发挥作用的具体机制,也为通过基因工程改良作物农艺性状提供了新的理论依据和靶点。
具体体现在以下几个方面:
揭示了QQS在紫花苜蓿中的作用机制: 研究首次发现并证实,源于模式植物的QQS基因能够与紫花苜蓿的内源核转录因子MsNF-YC4发生直接的蛋白质相互作用(Fig. 2A, 2B)。这解释了QQS作为一个非转录因子,是如何通过“借力”宿主自身的调控网络来影响下游基因表达和生理过程的,这是机制上的一个关键突破。
鉴定了MsNF-YC4的核心枢纽作用: 研究不仅证明了MsNF-YC4是连接QQS与下游信号的“桥梁”,还通过过表达实验(Fig. 3, 4)和分子生物学分析,清晰地阐明了MsNF-YC4自身作为一个响应干旱胁迫的核蛋白,在促进根系分枝和提升抗旱性中扮演着关键角色。
打通了从上游互作到下游靶基因的完整通路: 研究通过转录组学分析锁定了生长素信号通路(Fig. 5A),并进一步通过酵母单杂交和双荧光素酶报告实验,证实了MsNF-YC4能够直接结合到生长素响应因子MsARF8的启动子上并激活其转录(Fig. 6D, 6E)。这成功地将上游的蛋白互作与下游具体的基因调控事件联系起来,构建了一条清晰、线性的信号链。
验证了关键下游基因的功能: 通过反义寡核苷酸(asODN)技术对MsARF8进行瞬时抑制,实验有力地证明了MsARF8是生长素诱导的根系分枝和抗旱性的正向调控因子(Fig. 7)。这不仅为整个调控通路提供了功能性闭环证据,也凸显了MsARF8作为一个潜在的育种改良靶点的重要性。







 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
