2.1 植物材料培养、转基因株系构建与表型组织学分析
为探究目标基因的生物学功能,研究采用紫花苜蓿(‘中苜一号’)和拟南芥(Col-0)野生型作为试验材料,在标准温室和光照培养箱条件下进行培育。在转基因株系构建方面,将MsDUF3700的编码区序列克隆至表达载体中,随后通过农杆菌介导法(紫花苜蓿采用叶片外植体转化,拟南芥采用浸花法)获得过表达植株。为评估基因对植物生长的影响,对野生型和MsDUF3700过表达紫花苜蓿进行了系统的表型和组织化学分析,包括利用游标卡尺测量茎粗,并将茎节段经FAA固定、石蜡包埋和切片后,进行番红-固绿染色及显微镜观察,以明确其在细胞和组织水平的形态学变化。
2.2 基因表达定量、内源激素测定与蛋白质降解评估
为阐明MsDUF3700调控植物生长的生理与分子基础,研究提取了植物总RNA并反转录为cDNA,以MsActin为内参基因进行实时荧光定量PCR(RT-qPCR),从而精确评估相关基因的转录水平。在激素代谢层面,采用UPLC-MS/MS技术对野生型和MsDUF3700OE植株叶片中的内源赤霉素(GAs)含量进行了靶向提取与定量分析。此外,为探究DELLA蛋白的稳定性,设计了无细胞降解试验和体内免疫印迹分析:前者将纯化的GST-MsDELLA重组蛋白与植物总蛋白提取物共孵育(并设置蛋白酶体抑制剂MG132对照),后者则在本氏烟草叶片中瞬时共表达MsDUF3700-MYC与MsDELLA-GFP,随后通过SDS-PAGE分离和特异性抗体孵育,系统监测了DELLA蛋白在体外和体内的降解动态。
2.3 亚细胞定位、蛋白互作机制验证与统计学分析
为解析核心调控模块的分子互作网络,首先通过在本氏烟草中瞬时表达MsSLY1-GFP融合蛋白并利用共聚焦显微镜观察,确定了MsSLY1的亚细胞定位。随后,综合运用多种技术手段全面验证了MsDUF3700、MsSLY1与MsDELLA之间的物理相互作用:利用酵母双杂交(Y2H)试验检测了不同截短片段间的直接结合能力;通过双分子荧光互补(BiFC)和萤光素酶互补成像(LCI)试验在植物体内原位可视化了蛋白复合体的形成;并借助免疫共沉淀(Co-IP)试验进一步证实了蛋白间的体内结合。在数据处理方面,利用ImageJ软件测量形态学指标,所有试验均包含至少三个生物学重复,数据依托SPSS 27.0软件进行单因素方差分析(One-way ANOVA)以确定统计学显著性(P < 0.05),并使用GraphPad Prism软件完成最终的数据可视化。
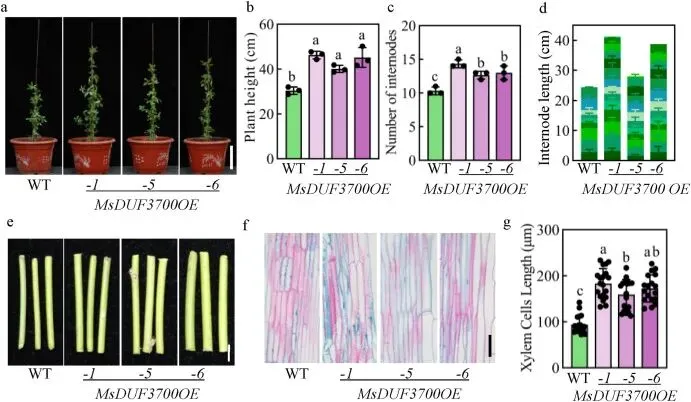
3.1 过表达MsDUF3700促进紫花苜蓿的植物生长和茎发育
基于前期全基因组关联分析鉴定出的候选基因MsDUF3700,对其过表达转基因株系(MsDUF3700OE)进行了全面的表型鉴定。结果表明,与野生型(WT)相比,过表达该基因显著增加了紫花苜蓿的株高、地上生物量和茎粗。形态学和组织学分析进一步揭示,株高的增加归因于节间数量的增多和节间的伸长,且转基因植株的木质部细胞显著长于野生型;同时,茎粗的增加伴随着髓腔的扩大和韧皮部组织的扩张。这些结果表明MsDUF3700协同促进了紫花苜蓿茎的纵向和径向扩展 (Fig. 1)。
图1. 过表达MsDUF3700促进紫花苜蓿的植物生长和茎发育。(a) 10周龄野生型 (WT) 和MsDUF3700过表达 (MsDUF3700OE) 转基因株系的表型。比例尺=10 cm。(b-d) 10周龄 WT 和 MsDUF3700OE 株系的株高 (b)、节间数 (c) 和节间距 (d) 的定量测量。节间长度通过测量从茎基部到顶端的单个节间来计算。(e) WT 和 MsDUF3700OE 株系从基部起第三节间的茎粗比较。比例尺 = 5 mm。(f) 显示 WT 和 MsDUF3700OE 株系细胞伸长的茎纵切面。比例尺=50 μm。(g) WT 和 MsDUF3700OE 植株木质部细胞长度的定量。不同小写字母表示差异有统计学意义 (P < 0.05,Duncan检验)。
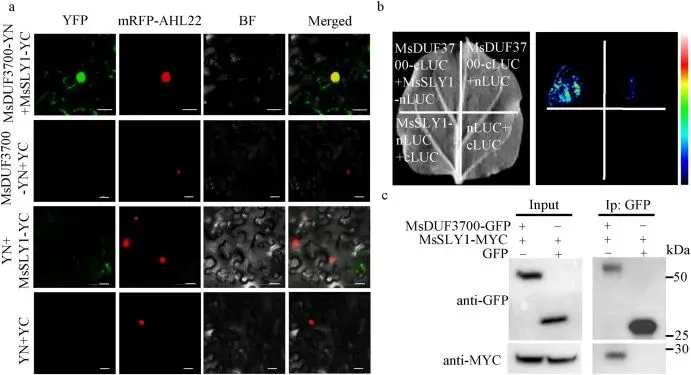
3.2 MsDUF3700与F-box蛋白MsSLY1发生物理相互作用
鉴于MsDUF3700定位于细胞核,为探究其促生长的分子机制,利用酵母双杂交(Y2H)筛选核cDNA文库,鉴定出一个与拟南芥赤霉素(GA)信号通路核心组分SLEEPY1 (SLY1) 高度同源的F-box蛋白,命名为MsSLY1。该蛋白主要在幼叶、根茎和花中表达,并定位于细胞核。随后,通过双分子荧光互补(BiFC)、萤光素酶互补成像(LCI)以及免疫共沉淀(Co-IP)等三种独立的试验方法,全面证实了MsDUF3700与MsSLY1在本氏烟草体内存在真实的物理相互作用,这为MsDUF3700与GA信号传导机制之间的功能联系提供了有力证据 (Fig. 2)。
图2. MsDUF3700与MsSLY1发生物理相互作用。(a) 双分子荧光互补 (BiFC) 试验显示 MsDUF3700 和 MsSLY1 在本氏烟草叶表皮细胞的细胞核内发生相互作用。AHL22 (At-hook基序核定位蛋白22) 用作核标记。比例尺=20 μm。(b) 萤光素酶互补成像 (LCI) 试验证实 MsDUF3700 和 MsSLY1 之间的相互作用。共表达 MsDUF3700-cLUC 和 MsSLY1-nLUC 产生强烈的发光信号。空载体或单一构建体用作阴性对照。(c) 免疫共沉淀 (Co-IP) 试验证实 MsDUF3700 和 MsSLY1 之间的相互作用。使用抗GFP抗体对表达 MsDUF3700-GFP 和 MsSLY1-MYC 的本氏烟草蛋白提取物进行免疫沉淀,并使用抗GFP和抗MYC抗体进行检测。
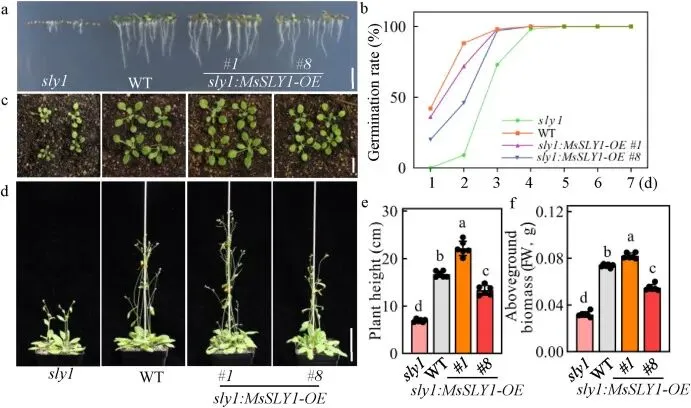
3.3 异位表达MsSLY1促进拟南芥的生长
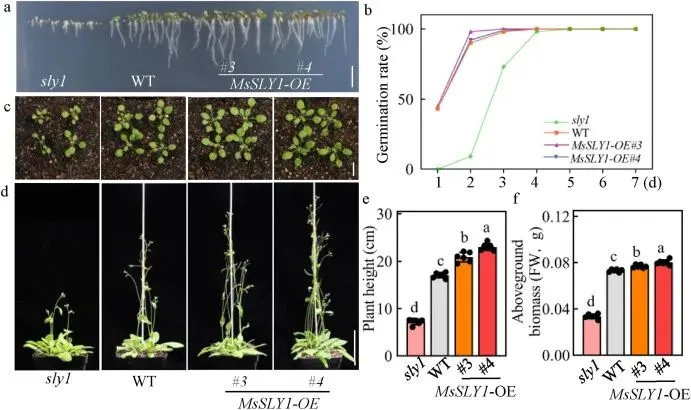
为进一步表征MsSLY1的功能,将其编码序列引入拟南芥sly1突变体中。结果显示,异位表达MsSLY1能够部分恢复sly1突变体的生长缺陷,包括恢复根系伸长、提高种子发芽率、促进子叶扩张以及增加株高和地上生物量,表明SLY1在物种间具有功能保守性 (Fig. 3)。此外,在野生型拟南芥背景下过表达MsSLY1,转基因株系同样表现出优于对照的生长性能,如主根更长、发芽更快、子叶面积更大以及株高和生物量增加。这些结果一致表明,MsSLY1在拟南芥中作为植物生长的保守正调控因子发挥作用 (Fig. 4)。
图3. 异位表达MsSLY1挽救了sly突变体的生长缺陷。(a, b) sly1、WT 和 sly1:MsSLY1-OE 株系的发芽表型 (a) 和7天后的发芽率 (b)。(a) 中拍摄了在 1/2 MS 培养基上生长的3日龄幼苗。比例尺=0.5 cm。(b) 中的数据以平均值表示 (n=3 个培养皿,每个培养皿接种100粒种子)。(c) 2周龄 sly1、WT 和 sly1:MsSLY1-OE 植株的莲座叶表型。比例尺=1 cm。(d-f) 4周龄植株的表型 (d)、株高 (e) 和地上生物量 (f)。比例尺=5 cm。数据为平均值±标准差 (n=6)。不同小写字母表示差异有统计学意义 (P < 0.05,Duncan检验)。
图4. 过表达MsSLY1促进野生型拟南芥的生长。(a, b) sly1、WT 和 MsSLY1-OE 转基因株系的发芽表型 (a) 和7天后的发芽率 (b)。比例尺=0.5 cm。(a) 中拍摄了在 1/2 MS 培养基上生长的3日龄幼苗。(b) 中的数据以平均值表示 (n=3 个培养皿,每个培养皿接种100粒种子)。(c) 2周龄 sly1 突变体、WT 和 MsSLY1-OE 植株的莲座叶表型。比例尺=1 cm。(d-f) 4周龄 sly1 突变体、WT 和 MsSLY1-OE 植株的表型 (d)、株高 (e) 和地上生物量 (f)。比例尺=5 cm。数据为平均值±标准差 (n=6)。不同小写字母表示差异有统计学意义 (P < 0.05,Duncan检验)。
3.4 MsSLY1与MsDELLA互作并促进其降解
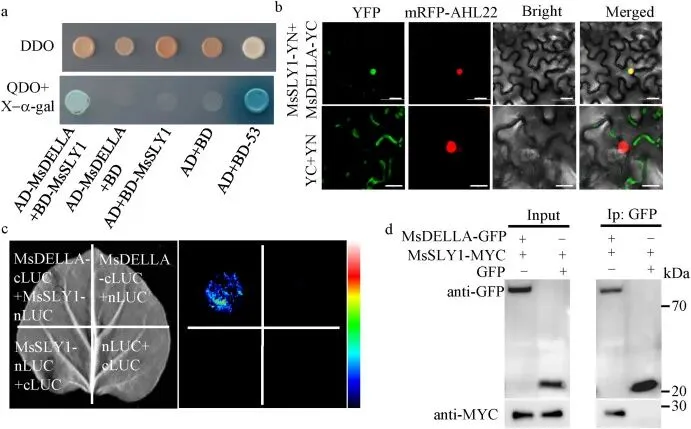
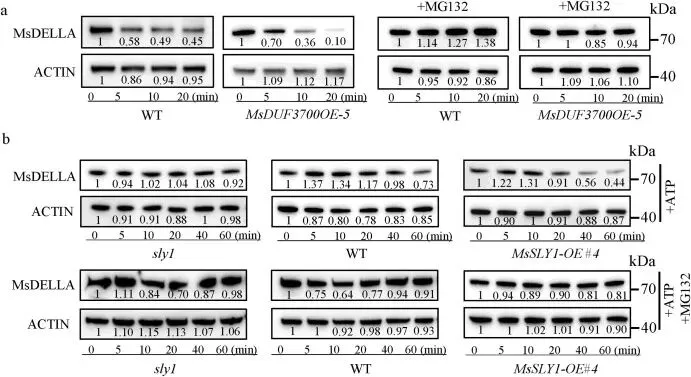
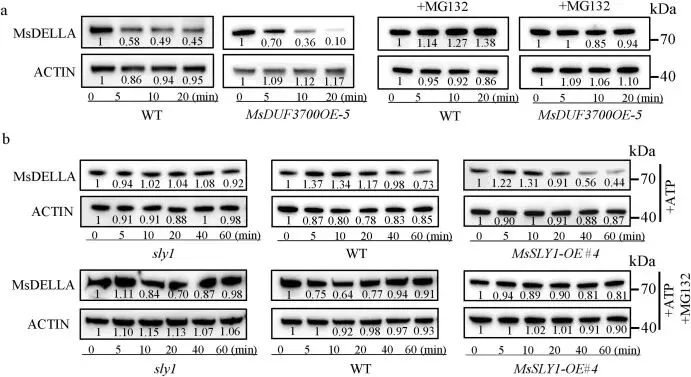
鉴于SLY1在拟南芥中能靶向降解DELLA蛋白,研究进一步检验了MsSLY1是否与紫花苜蓿GA信号关键负调控因子MsDELLA发生互作。Y2H、BiFC、LCI和Co-IP等试验一致证实了全长MsSLY1与MsDELLA在体内外的物理互作,且MsSLY1的C端结构域是识别MsDELLA所必需的 (Fig. 5)。为评估这种互作对MsDELLA稳定性的影响,无细胞降解试验表明,在缺乏蛋白酶体抑制剂MG132的情况下,MsDELLA在MsDUF3700OE紫花苜蓿提取物中的降解速度快于野生型,而MG132处理可阻断该降解过程;拟南芥提取物试验也得出了类似结论。此外,植物体内瞬时共表达分析显示,MsDUF3700的加入会导致MsDELLA蛋白水平明显降低。这些结果共同表明,MsDUF3700通过泛素-蛋白酶体途径促进MsSLY1介导的DELLA降解,从而调节GA信号输出 (Fig. 6)。
图5. MsSLY1与MsDELLA发生物理相互作用。(a) 酵母双杂交 (Y2H) 试验显示 MsSLY1 和 MsDELLA 之间的相互作用。AD,激活结构域;BD,结合结构域;DDO,SD/-Trp/-Leu;QDO,SD/-Trp/-Leu/-His/-Ade/X-a-gal。(b) BiFC 试验显示 MsSLY1-YN 和 MsDELLA-YC 在本氏烟草叶细胞的细胞核内发生相互作用。AHL22 用作核标记。比例尺=20 μm。(c) LCI 试验显示 MsSLY1 和 MsDELLA 之间发出强烈的发光信号。空载体或单一构建体用作阴性对照。色标代表 LUC 活性。(d) Co-IP 试验证实了本氏烟草中 MsSLY1-MsDELLA 的相互作用。使用抗GFP抗体对蛋白提取物进行免疫沉淀,并使用抗GFP和抗MYC抗体进行分析。
图6. MsSLY1通过泛素-蛋白酶体途径促进MsDELLA降解。(a) 使用来自 WT 和 MsDUF3700OE 紫花苜蓿的总蛋白与带有 GST 标签的 MsDELLA 孵育进行的无细胞降解试验。MG132 用作蛋白酶体抑制剂。(b) 使用来自1周龄 WT、sly1 和 MsSLY1-OE 拟南芥的提取物降解带有 GST 标签的 MsDELLA。对蛋白丰度进行定量并标准化至 0 分钟时间点。
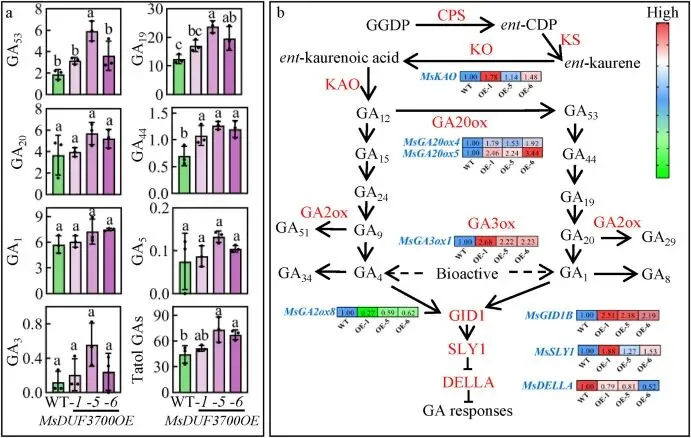
3.5 MsDUF3700调节赤霉素的生物合成与信号传导
为探究MsDUF3700促生长的内分泌机制,对内源GA代谢物进行了靶向定量分析。结果显示,MsDUF3700OE植株中GA合成前体(GA53、GA44和GA19)的积累量显著高于野生型,而生物活性GA的变化较为温和,表明MsDUF3700主要影响GA代谢通量而非诱导活性激素的过度积累 (Fig. 7a)。与此代谢转变相一致,RT-qPCR分析揭示了关键GA合成基因(MsKAO1/2、MsGA20ox4/5、MsGA3ox1)在转基因植株中显著上调,而GA失活基因(MsGA2ox8)被下调。在信号传导层面,GA受体MsGID1B和F-box蛋白MsSLY1的表达上调,而信号抑制因子MsDELLA的转录本下调。这些结果表明,MsDUF3700通过在多个调控层面上协调改变GA代谢谱和重编程关键基因的转录,从而微调GA通路活性以促进紫花苜蓿的茎伸长和生物量积累 (Fig. 7)。
图7. MsDUF3700调节紫花苜蓿中GA的生物合成和信号传导。(a) 紫花苜蓿10周龄 WT 和 MsDUF3700OE 转基因株系叶片中内源中间体 GA 含量 (ng/g)。数据为平均值±标准差 (n=3)。(b) 基于 RT-qPCR 分析显示 GA 生物合成和信号传导基因相对转录水平的热图。WT 中的表达水平标准化为 1。不同小写字母表示差异有统计学意义 (P < 0.05,Duncan检验)。
3.6 MsDUF3700与MsSLY1的遗传关系
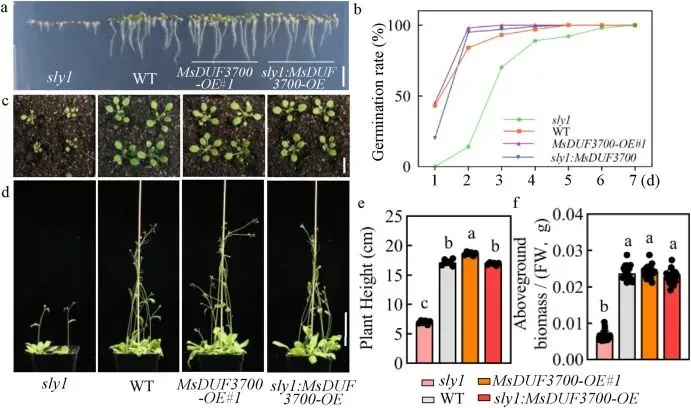
为深入探讨MsDUF3700与MsSLY1之间的遗传上位性,研究在拟南芥突变体和野生型背景下构建了MsDUF3700过表达株系。在野生型拟南芥中异位表达MsDUF3700仅导致株高适度且具有统计学意义的增加,而在发芽或茎粗上无明显差异,暗示该基因单独作用不足以显著改变拟南芥的生长。进一步将MsDUF3700在sly1突变体背景下过表达(sly1:MsDUF3700-OE),发现其能部分挽救sly1突变特征性的矮化和GA不敏感表型,包括改善根长、发芽率、子叶大小以及后期的株高、生物量和茎粗。然而,这种表型挽救是不完全的,表明MsDUF3700不能完全补偿SLY1功能的缺失。这些遗传互作证据支持了一种模型,即MsDUF3700在MsSLY1上游或与其协同作用以调节DELLA周转和GA信号输出,而不是绕过经典的依赖于SLY1的途径 (Fig. 8)。
图8. 拟南芥中DUF3700和SLY1的遗传上位性分析。(a, b) sly1、WT、MsDUF3700-OE 和 sly1:MsDUF3700-OE 株系的发芽表型 (a) 和7天后的发芽率 (b)。(a) 中拍摄了在 1/2 MS 培养基上生长的3日龄幼苗,并在 (b) 中对7日龄幼苗进行了统计分析。(b) 中的数据以平均值表示 (n=3 个培养皿,每个培养皿接种100粒种子)。(c) 2周龄 sly1、WT、MsDUF3700-OE 和 sly1:MsDUF3700-OE 植株的莲座叶表型。(d-f) 4周龄 sly1、WT、MsDUF3700-OE 和 sly1:MsDUF3700-OE 植株的表型 (d)、株高 (e) 和地上生物量 (f)。(a) 中比例尺=5 cm,(c) 中为1 cm,(d) 中为5 cm。数据为平均值±标准差 (n=6)。不同小写字母表示差异有统计学意义 (P < 0.05,Duncan检验)。







 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
