温度是影响植物生长发育的关键环境因子,热胁迫可导致光合抑制、叶片衰老甚至植株死亡。在全球气候变暖背景下,解析植物耐热机制具有重要理论与应用价值。研究表明,DNA甲基化可通过调控启动子活性影响胁迫相关基因表达。尽管在拟南芥、棉花、甘蓝等物种中已有相关发现,但作为喜凉叶菜作物的不结球白菜(NHCC)对热胁迫高度敏感,其耐热性的表观遗传调控机制仍不清楚。因此,揭示DNA甲基化在NHCC热胁迫响应中的作用,对理解其耐热机理及分子育种具有重要意义。
2026年1月7日,上海市农业科学院朱红芳团队联合上海交通大学方玉达教授在Journal of Agricultural and Food Chemistry上发表题为“Integrative Analysis of DNA Methylome and Transcriptome Reveals the Mechanism of Heat Tolerance in Non-heading Chinese Cabbage”的研究论文,研究通过整合分析热胁迫下NHCC的DNA甲基化组和转录组数据,揭示了DNA高甲基化在NHCC耐热性调控中的正向作用。
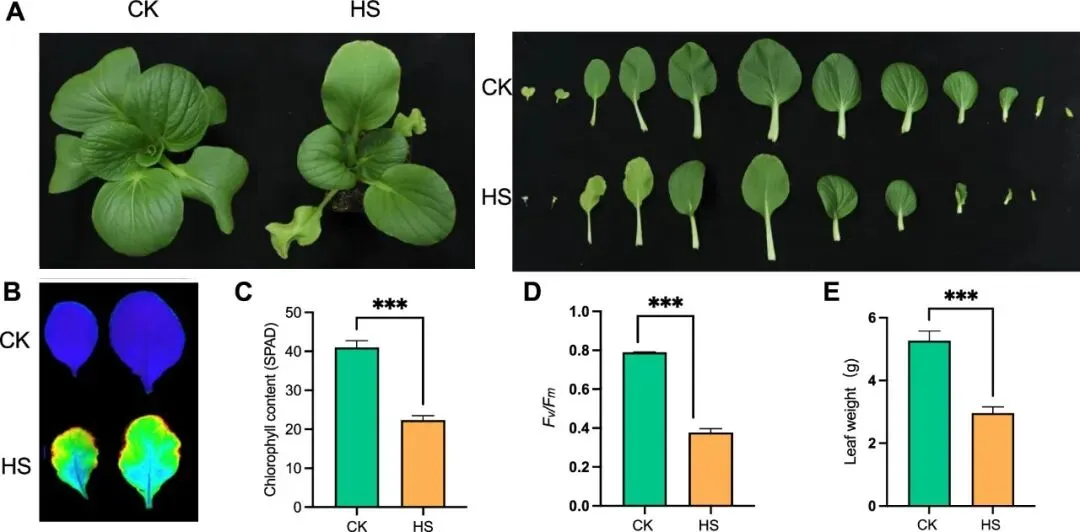
研究首先对2月龄的NHCC进行42℃、24h热胁迫处理,发现其叶片严重萎蔫(图1A)。生理指标显示,热胁迫显著促进叶绿素降解、降低光系统II最大光化学效率(Fv/Fm),并加速叶片失水(图1B–E),表明高温显著抑制NHCC的光合能力和水分保持,对其生长发育造成明显不利影响。
图1. 热应激处理破坏了NHCC的表型形态
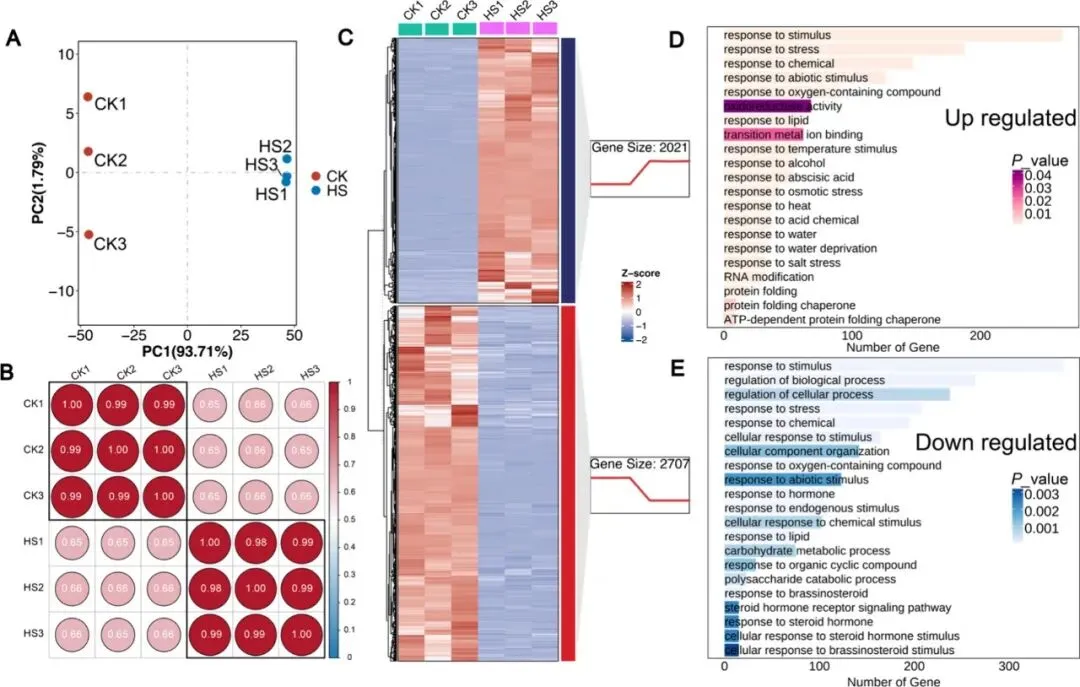
为揭示热胁迫影响NHCC的分子机制,研究对对照组与热胁迫处理组叶片进行RNA-seq分析。PCA和皮尔逊相关性分析显示样本重复性良好。以|log2FC| ≥ 2且P < 0.01为阈值,共鉴定出4728个差异表达基因(DEGs),其中2707个下调,2021个上调。GO富集分析表明,这些DEGs显著富集于热胁迫响应、非生物胁迫应答、温度感知、激素信号转导、水分缺失响应及初级代谢等通路,说明热胁迫通过大规模重编程转录组,干扰关键生理过程,进而引发NHCC的表型损伤。
图2. NHCC热胁迫诱导转录组分析
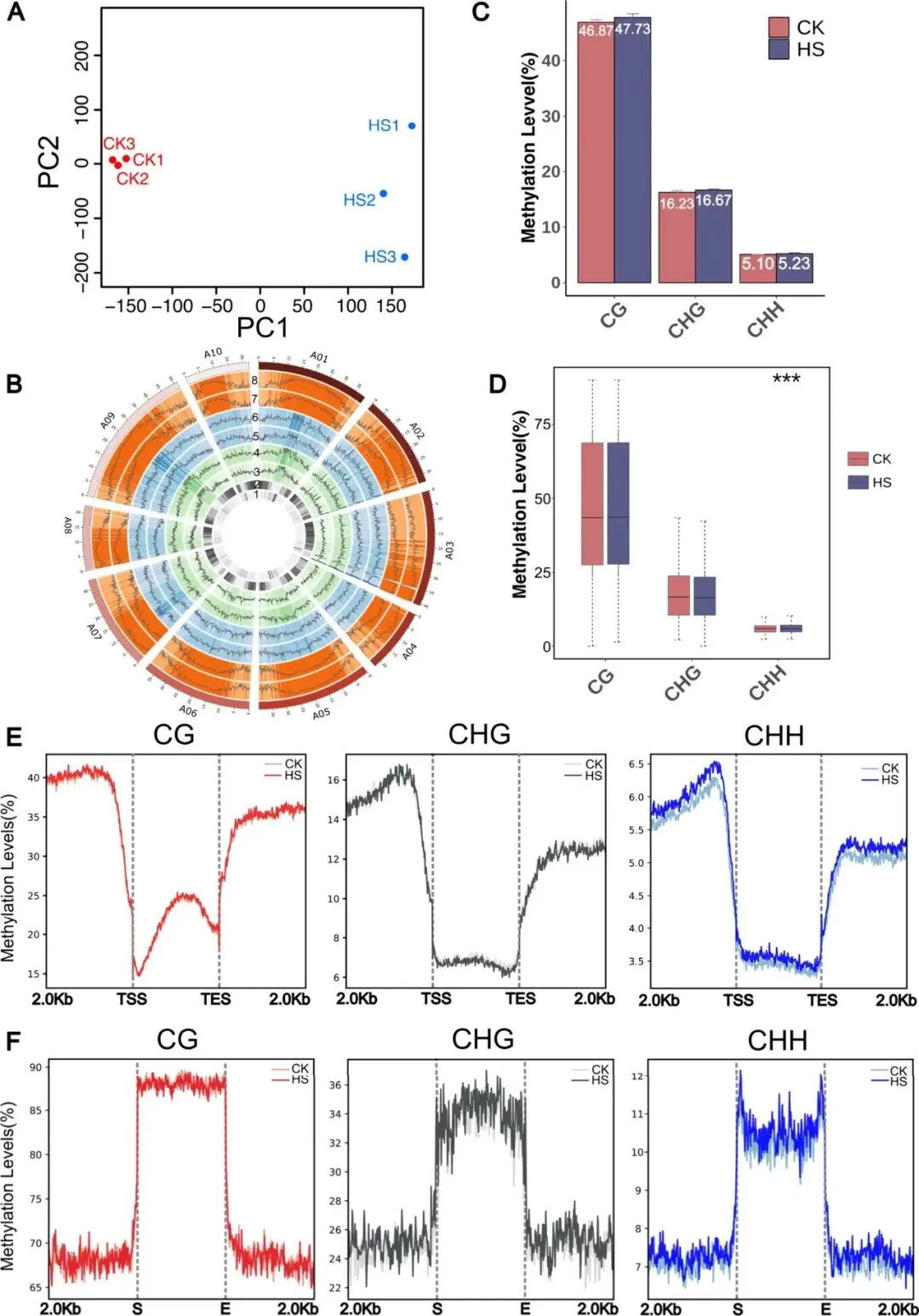
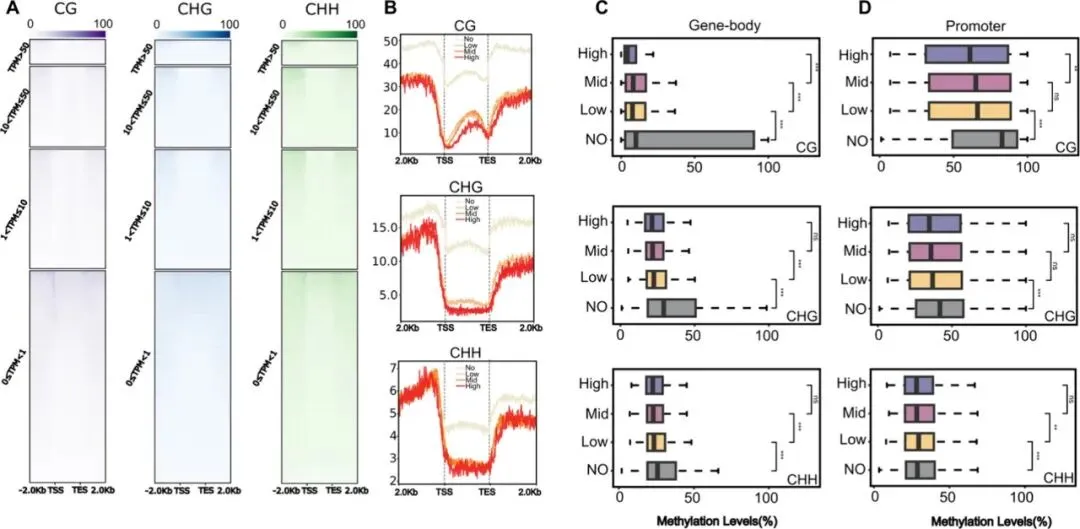
为深入解析DNA甲基化在NHCC热胁迫响应中的作用机制,研究采用WGBS进行分析。PCA显示样本重复性良好;全基因组特征表明,DNA甲基化水平与基因密度呈负相关,而与转座元件(TE)覆盖率呈正相关。热胁迫显著提升CHH甲基化水平,尤其在转录起始位点(TSS)上游2 kb的启动子区域增幅最为明显。此外,在占基因组约50%的TE区域,热胁迫主要诱导CHG和CHH甲基化变化。这些结果提示,启动子区CHH甲基化的动态调控可能在NHCC热胁迫适应中发挥关键作用。
图3. NHCC热应激对基因组甲基化影响的分析
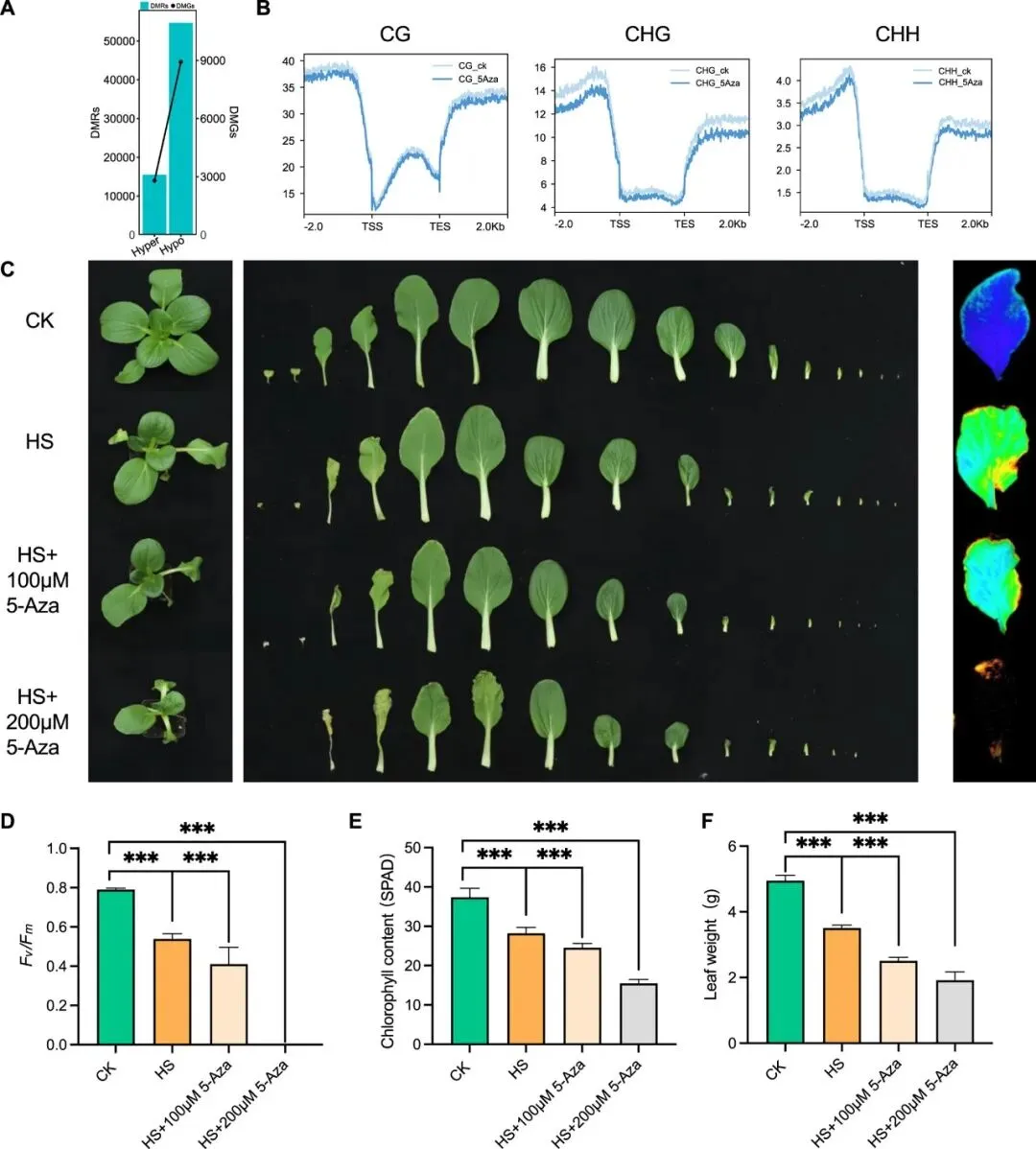
为验证DNA甲基化在NHCC耐热性中的功能,研究采用甲基化抑制剂5-氮杂胞苷(5-Aza)处理植株。结果显示,5-Aza显著降低全基因组CG、CHG和CHH序列的甲基化水平;经热胁迫后,处理组植株表现出更严重的叶片萎蔫和热损伤,且损伤程度随5-Aza浓度升高而加剧。生理指标显示,5-Aza处理显著降低光系统II最大光化学效率(Fv/Fm)、加速叶绿素降解并减少叶片鲜重。这些结果表明,DNA甲基化在NHCC热胁迫响应中发挥保护性调控作用,其水平下降会显著削弱植株耐热性。
图4. 检验5-Aza处理对NHCC耐热性的影响
为探究DNA甲基化对基因表达的调控作用,研究基于TPM值将NHCC全基因组基因分为四组进行分析。结果显示,在CG、CHG和CHH三种序列背景下,DNA甲基化水平均与基因表达呈显著负相关:转录沉默或低表达基因(0 ≤ TPM < 1)的甲基化水平最高。进一步分析基因本体及其侧翼区域发现,尽管高、低表达基因间甲基化差异有限,但沉默基因在所有序列背景下均呈现最强甲基化信号。对基因区及启动子区的统计分析再次证实,甲基化程度越高,转录活性越低。上述结果表明,DNA甲基化在NHCC中主要发挥转录抑制作用。
图5. DNA甲基化与基因表达的关系分析
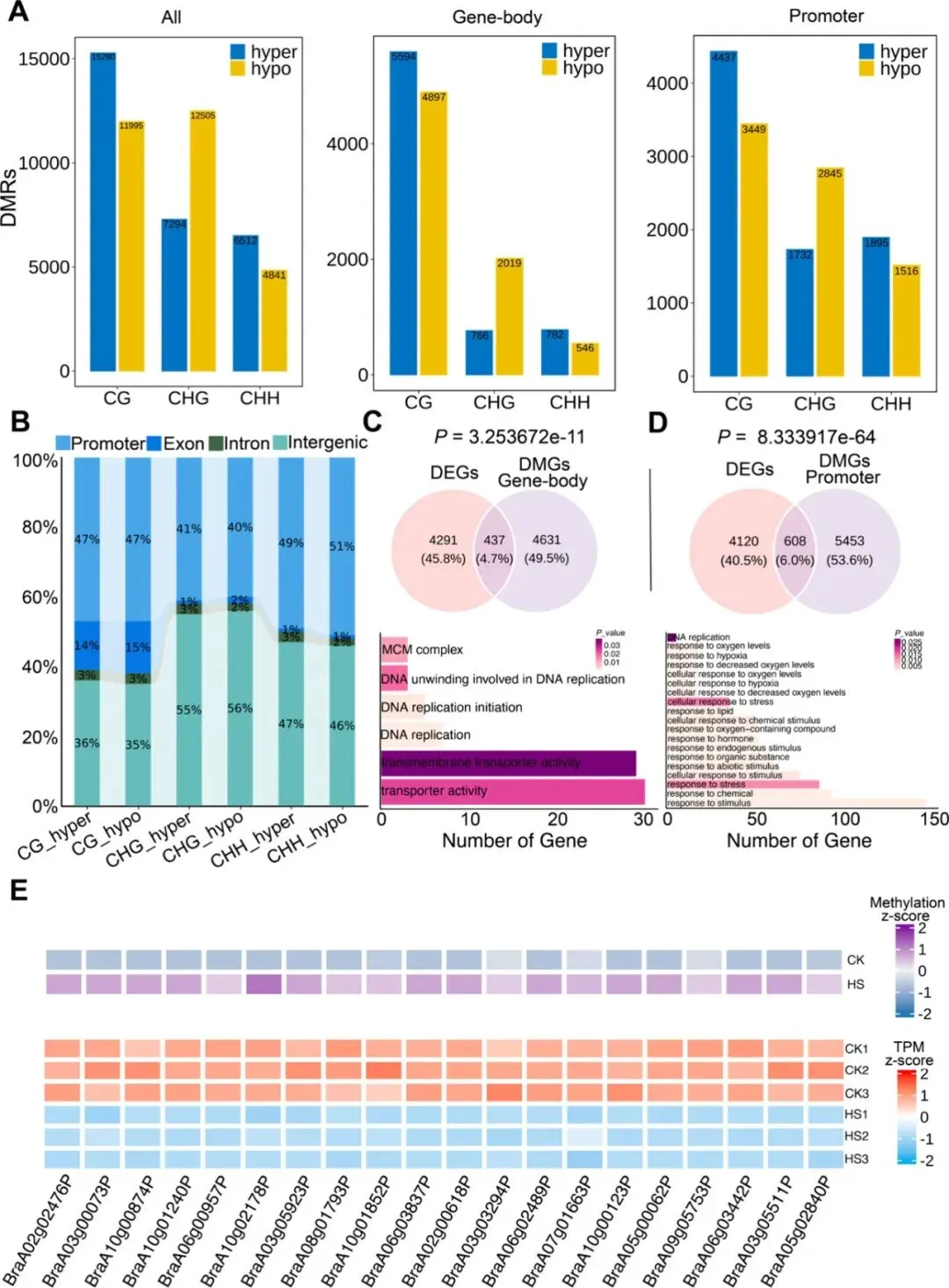
为解析热胁迫下DNA甲基化与基因表达的关联,研究鉴定出58457个差异甲基化区域(DMRs),其中CG和CHH类型的甲基化变化最为显著,且主要富集于启动子区。GO分析显示,不同序列背景(CG、CHG、CHH)的DMRs关联的功能通路各异;尤其值得注意的是,启动子区发生甲基化变化的DEGs显著富集于刺激响应和胁迫响应通路,而基因体甲基化相关的DEGs则多参与DNA复制与转运过程。基于此,研究进一步整合启动子高甲基化的差异甲基化基因(hyper-DMGs)与热胁迫下被抑制的DEGs,筛选出一批可能负调控NHCC耐热性的候选因子,为后续功能验证提供关键靶点。
图6. 热应激调节的DMGs和DEGs的综合分析
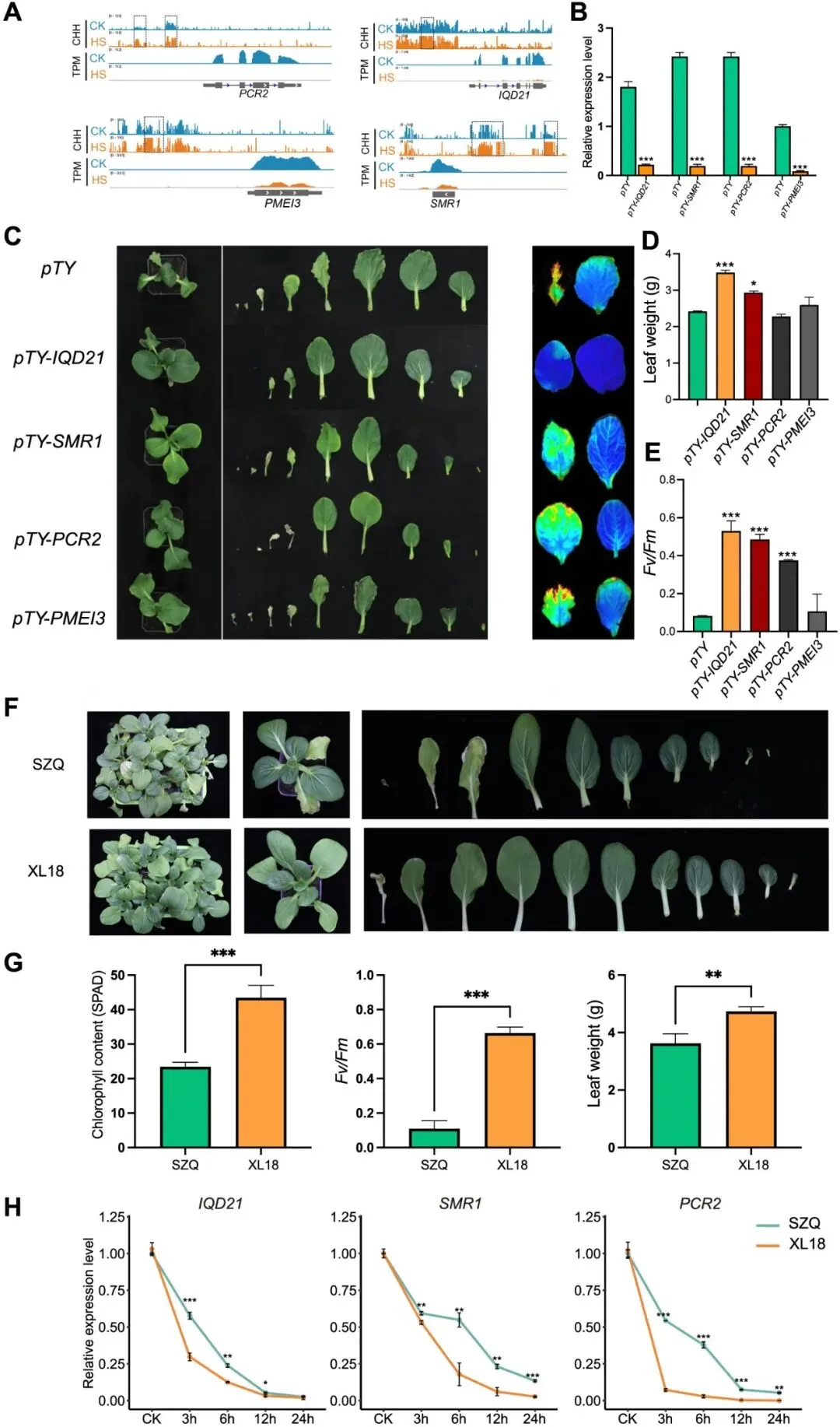
为验证候选基因在NHCC耐热性中的功能,研究结合甲基化与表达分析发现,IQD21、SMR1、PCR2和PMEI3启动子区CHH甲基化水平在热胁迫下显著升高,且其转录被强烈抑制。通过VIGS沉默实验表明,沉默IQD21、SMR1或PCR2可增强植株耐热性,表现为地上部生物量和/或光化学效率提升;而PMEI3沉默无显著效应。进一步比较耐热品种夏龙18号(XL18)与敏感品种苏州青(SZQ)发现,XL18在热胁迫下不仅表型更优,且IQD21、SMR1和PCR2的转录抑制更迅速、更显著。这些结果表明,这三个基因作为热胁迫负调控因子,其表达水平与NHCC耐热性呈负相关。
图7. NHCC热应激时下调的超DMGs热调节功能的检验
综上,这项研究揭示了DNA甲基化在NHCC适应热应激中的重要作用。热应激通过增加某些负调控热应激基因启动子区域的CHH甲基化,抑制其表达,从而提高NHCC的热耐受性。这些发现不仅揭示了NHCC应对热应激的分子机制,还为未来培育耐热性NHCC品种提供了重要的科学信息。

 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
