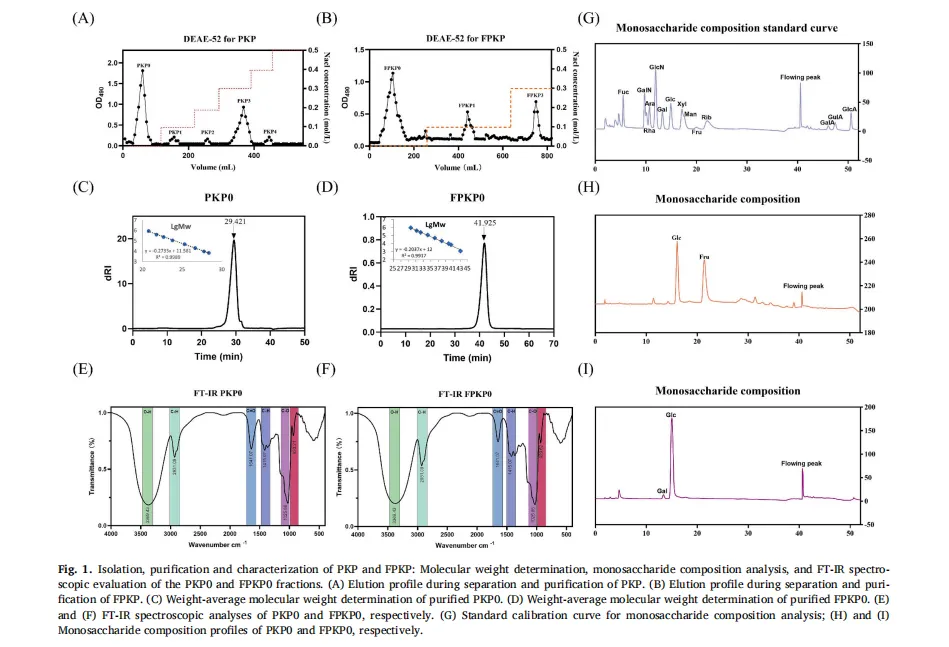
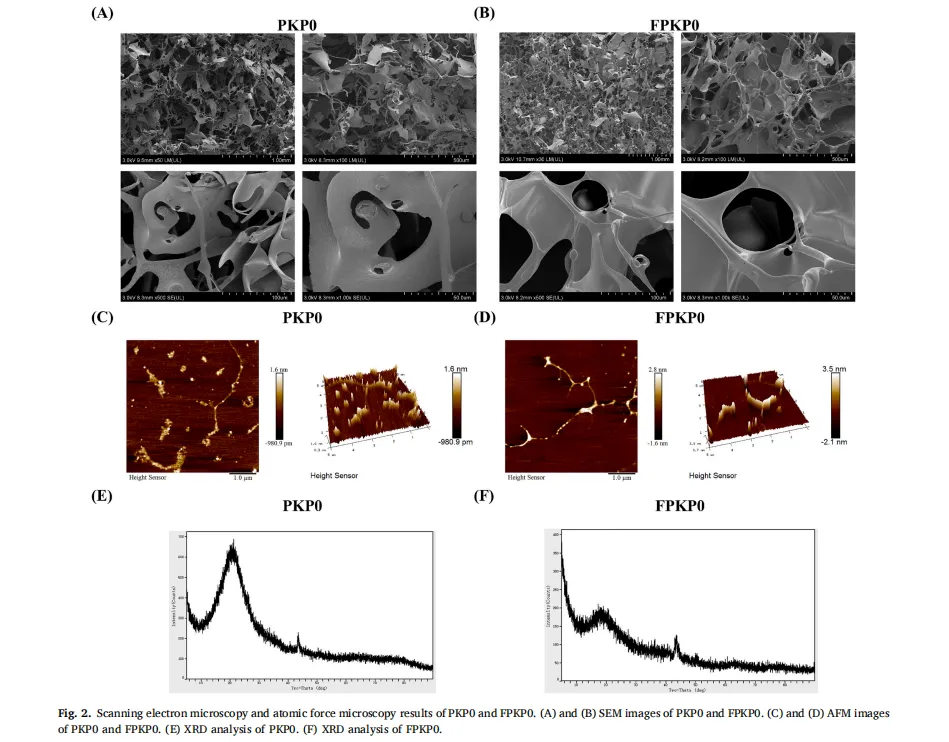
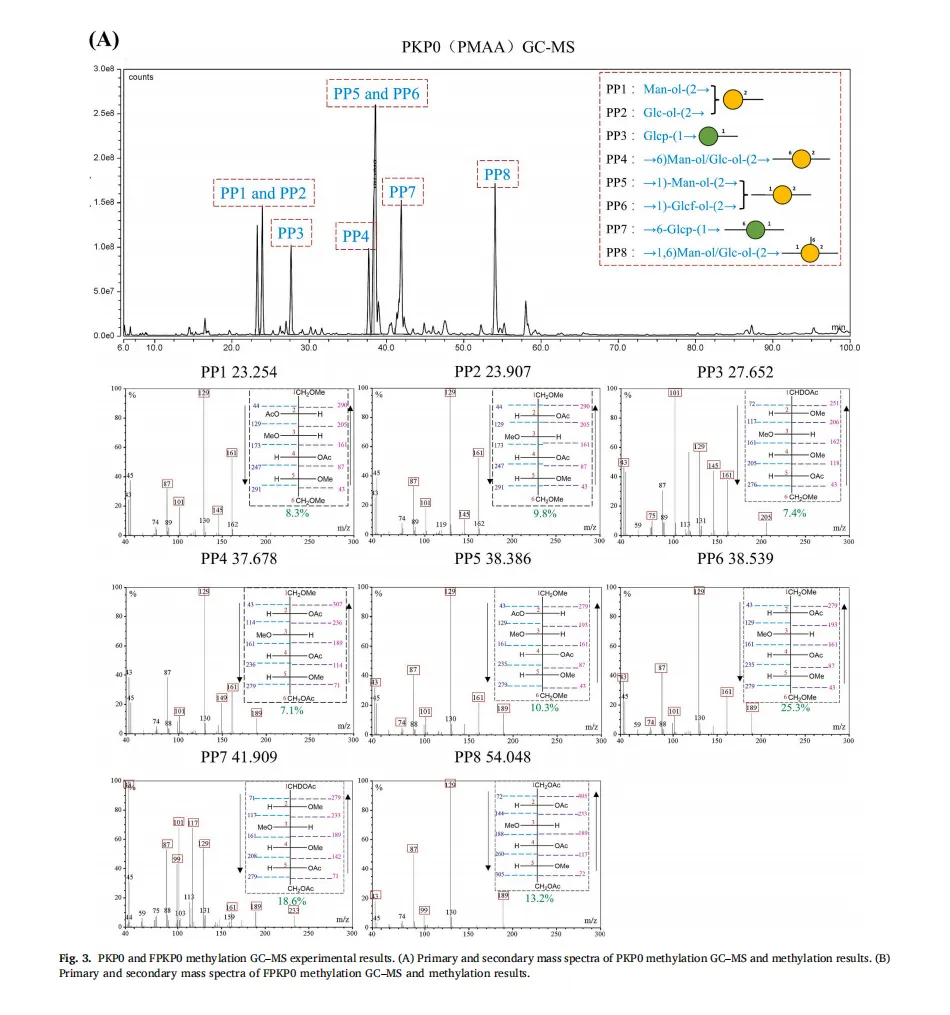
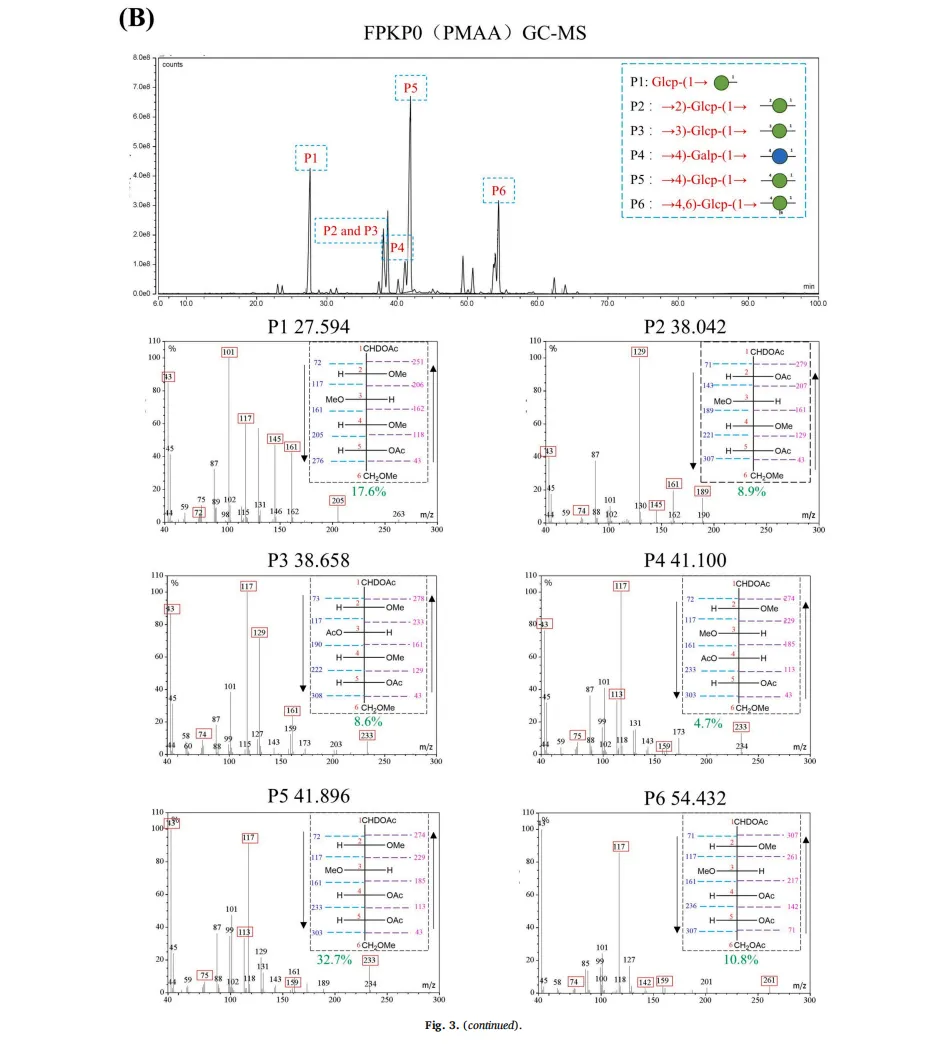
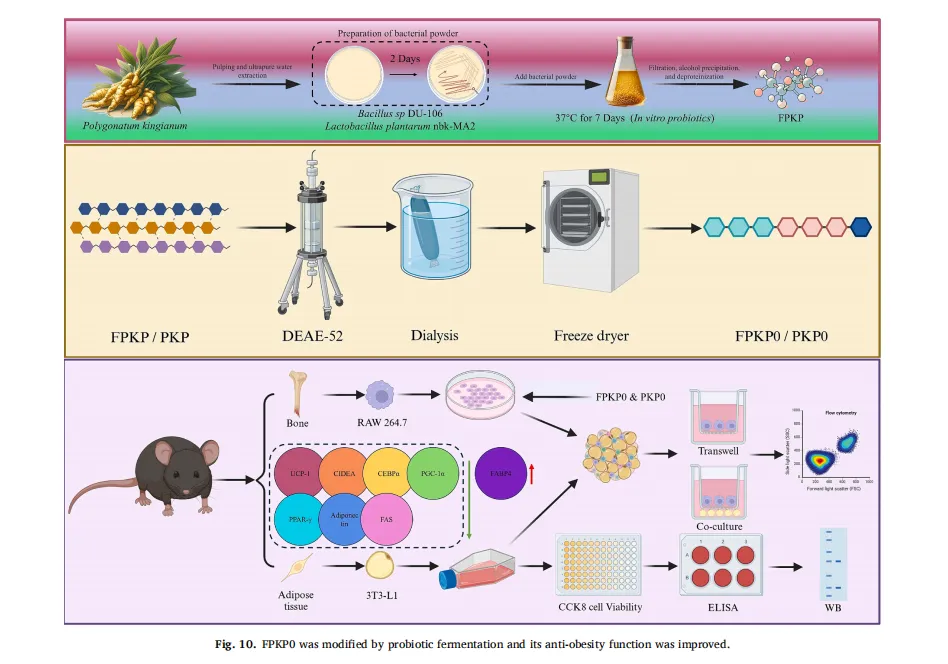
植物多糖通常存在于植物细胞壁、海藻和微生物大分子中。由于其天然来源、低毒性以及诸如抗炎、抗肥胖和促进脂质代谢作用的生物活性,植物多糖近年来引起了显著的研究关注。然而,传统的植物来源的多糖,如来自芦笋的菊粉型多糖,类似于膳食纤维,并且难以被人体吸收,这限制了它们的生物活性和实际应用。因此,人们对开发有效和可持续的生物转化方法以提高这些多糖的生物活性越来越感兴趣。
近年来,益生菌发酵已成为提高植物多糖生物活性的有效和可持续的方法。在发酵过程中,益生菌分泌酶,如糖苷酶和纤维素酶,其选择性地水解糖苷键,降低分子量并改变多糖的结构。这种结构修饰不仅提高了多糖的水溶性和生物利用度,而且通过暴露更多的活性基团或形成新的功能片段来增强其生物活性。益生菌发酵已被证明可以将三叶青多糖的分子量从1.82×104Da降低到1.23×104 Da,增加分支并暴露更多的活性基团,从而显着增加其抗氧化和免疫调节活性。类似地,苦瓜多糖的发酵部分水解β-糖苷键,产生更有效地与肠道受体结合的低分子量片段,从而增强抗肥胖活性。此外,豆瓣菜种子多糖经过发酵,破坏α-1,6-糖苷键,产生具有改善的抗糖尿病作用的生物活性低聚糖。然而,尽管这些研究证明了益生菌发酵在修饰多糖结构和增强其生物活性方面的潜力,但关于具体机制的许多问题仍未得到解答,例如发酵的影响是否在不同的多糖中有所不同以及分子量和分支结构的变化如何影响生物机制。这些方面仍需进一步调查。
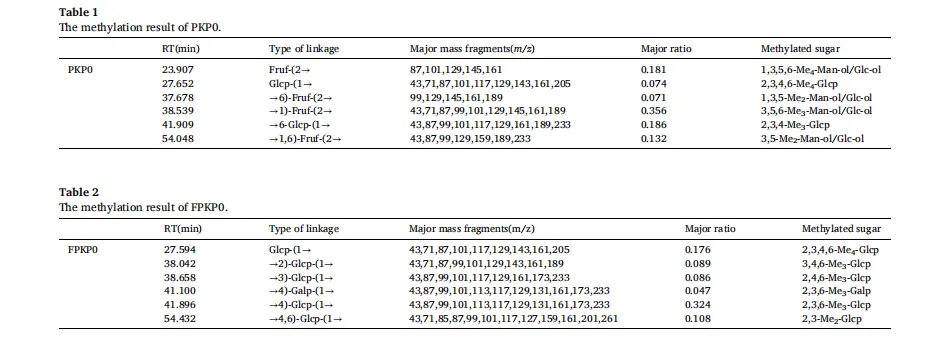
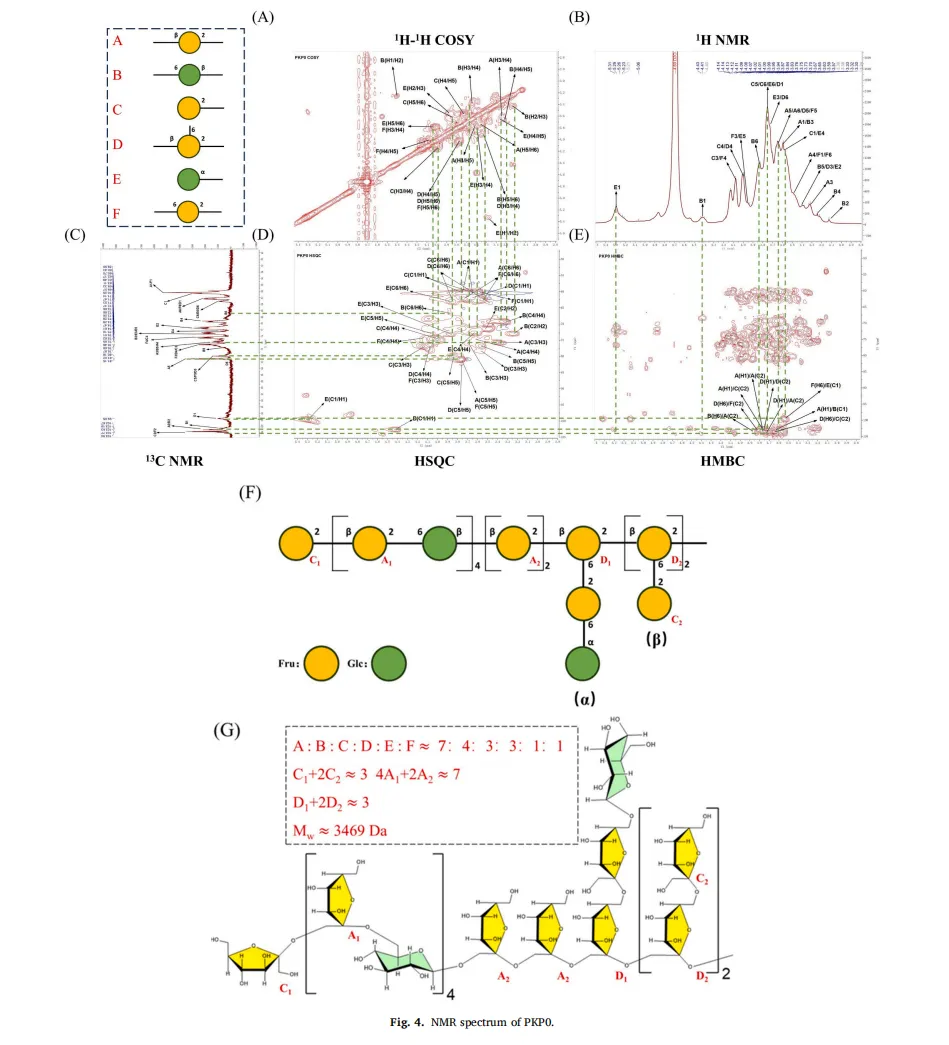
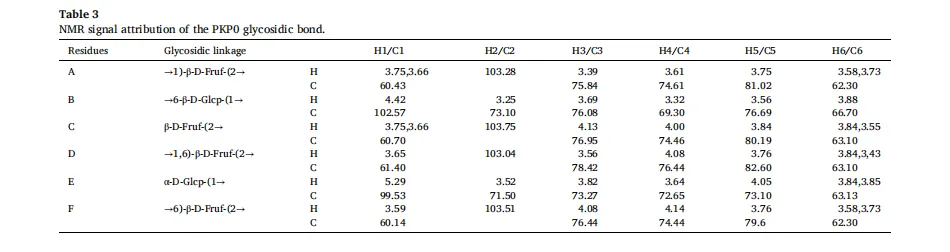
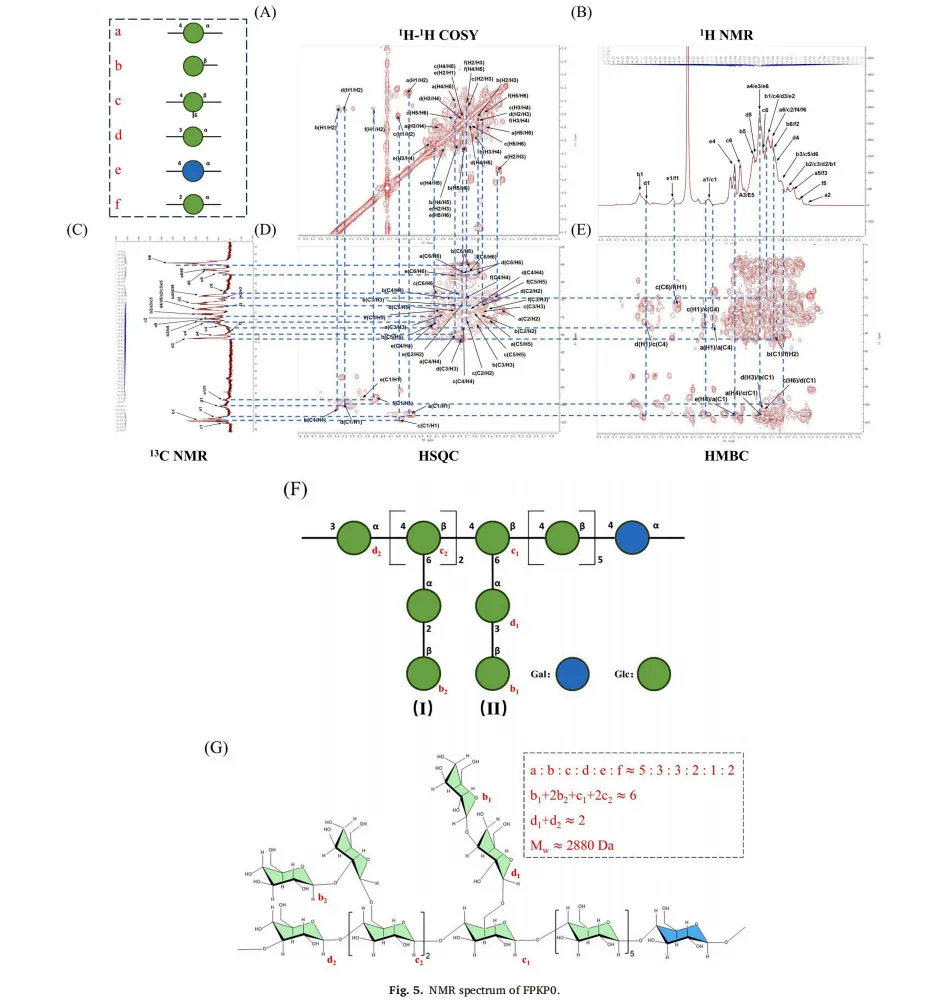
黄精(Polygonatum sibiricum(Hemsl))是我国传统的食药植物,主要分布于云南、湖南和东北地区。它已被广泛用于治疗肥胖,气血失调和代谢失衡。以前的研究已经确定多糖是主要的生物活性成分,负责其健康益处,特别是在减轻代谢疾病。例如,来自黄精的多糖已被证明可有效减轻高脂饮食诱导的小鼠肥胖,证明了显著的抗肥胖潜力。鉴于不同黄精属植物多糖结构的多样性导致其生物活性的差异,本研究以黄精属植物滇黄精多糖(Polygonatum kingianum,PKP)为研究对象。我们的初步实验结果表明,PKP也表现出显着的减肥作用。基于先前对来自各种来源的多糖的益生菌发酵的研究,假设益生菌发酵可以进一步改善PKP的抗肥胖活性,据我们所知,这是一个尚未充分探索的领域。鉴于PKP的单糖组成相对简单,其特征在于高比例(超过50%→1)-β-D-Fruf-(2→),如所报道的,研究益生菌发酵对PKP的结构和抗肥胖活性的影响将提供对益生菌发酵通过结构修饰增强其抗肥胖作用的机制的见解。