《食品科学》:南京林业大学勇强教授等:体外法分析不同肠段微生物对半乳甘露聚糖的利用规律
- 2026-07-12 08:35:13

01
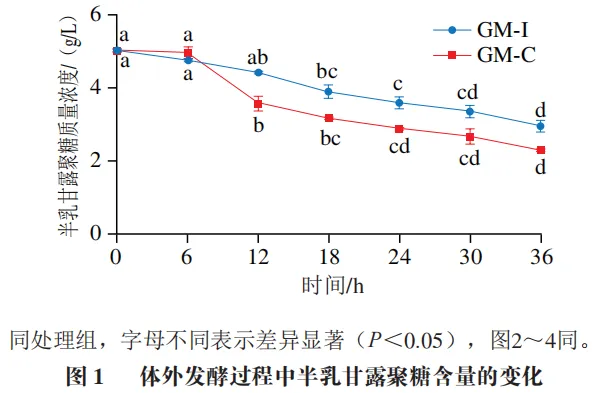
发酵过程中半乳甘露聚糖含量的变化
如图1所示,回肠和结肠发酵液中半乳甘露聚糖含量在发酵0~6 h后均无显著变化。回肠发酵液中半乳甘露聚糖含量在发酵18 h后显著降低(P<0.05),结肠发酵液中半乳甘露聚糖含量在发酵12 h后显著降低(P<0.05)。就利用能力而言,结肠微生物在发酵期间对半乳甘露聚糖的消耗量高于回肠微生物;就利用效率而言,回肠微生物对半乳甘露聚糖的消耗速率在整个发酵期较为一致,而结肠微生物对半乳甘露聚糖的消耗速率在发酵6~12 h时较高,其他时间段较低。
02
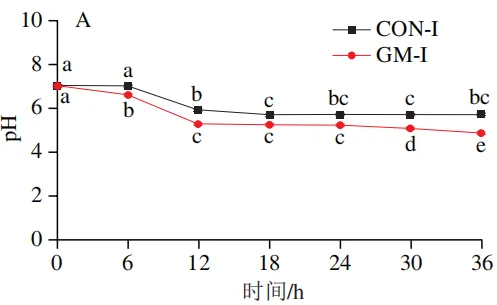
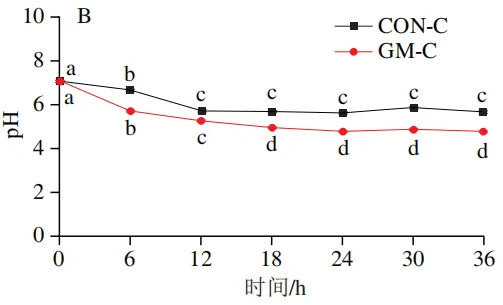
发酵过程中pH值的变化
如图2A所示,回肠对照组的pH值在发酵12 h和18 h后显著下降(P<0.05),随后无显著变化;回肠半乳甘露聚糖组的pH值在整个发酵期持续下降,且始终低于回肠对照组,其中发酵6、12、30 h和36 h时pH值的下降达到显著水平(P<0.05)。如图2B所示,结肠对照组的pH值在发酵6 h和12 h后显著下降(P<0.05),随后无显著变化;结肠半乳甘露聚糖组的pH值在发酵6、12 h和18 h后显著下降(P<0.05),随后趋于平缓,且结肠半乳甘露聚糖组pH值始终低于结肠对照组。
 |  |

03
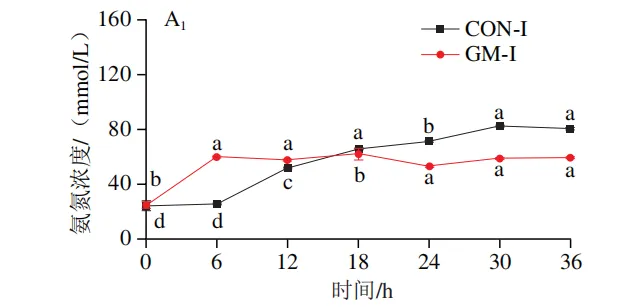
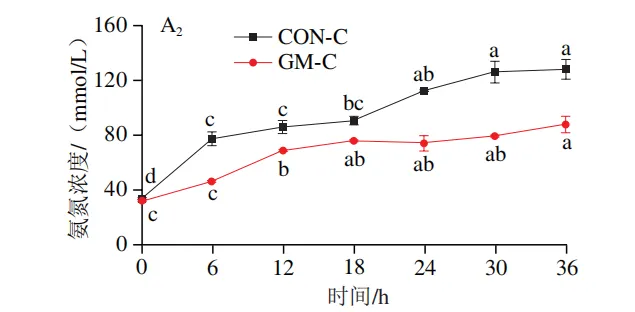
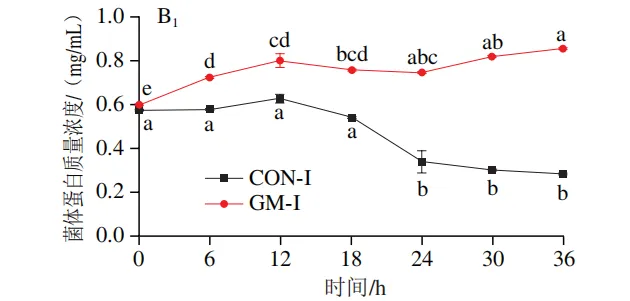
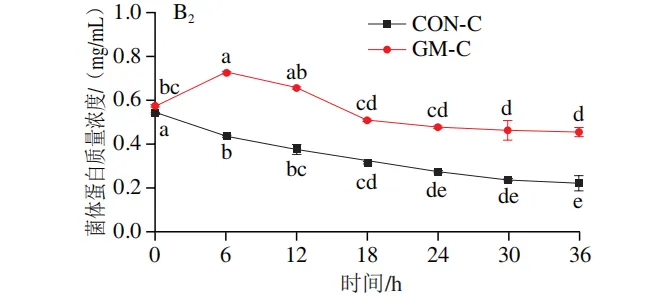
发酵过程中氨氮和菌体蛋白含量的变化
如图3A1、B1所示,在回肠发酵液中,对照组氨氮含量在发酵0~6 h后无显著变化,在发酵12~30 h后逐渐上升,在发酵30 h后趋于平稳;菌体蛋白含量在发酵0~18 h无显著变化,随后持续降低。半乳甘露聚糖组氨氮含量在发酵6 h时显著上升(P<0.05),随后无显著变化;菌体蛋白含量在发酵0~12 h持续升高,随后趋于平缓。如图3A2、B2所示,在结肠发酵液中,对照组氨氮含量在发酵0~30 h持续上升,随后趋于平缓;菌体蛋白含量在整个发酵期持续下降。半乳甘露聚糖组氨氮含量在发酵6~18 h持续上升,随后无显著变化,但始终低于结肠对照组;菌体蛋白含量在发酵0~6 h显著上升,在发酵6~36 h略有下降,但始终高于结肠对照组。
 |  |
 |  |

04
发酵过程中乳酸和短链脂肪酸含量的变化
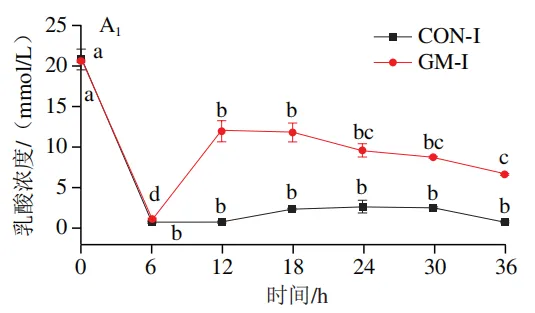
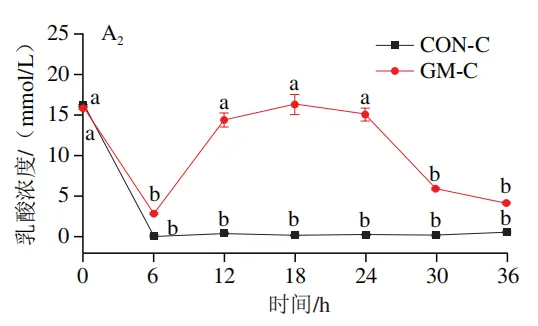
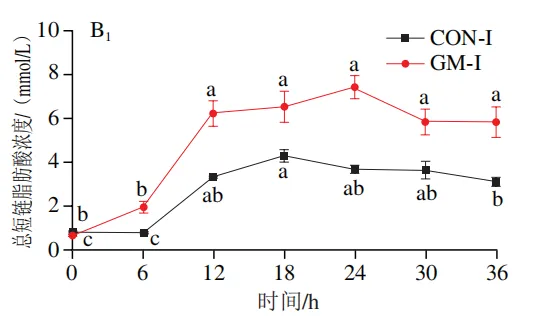
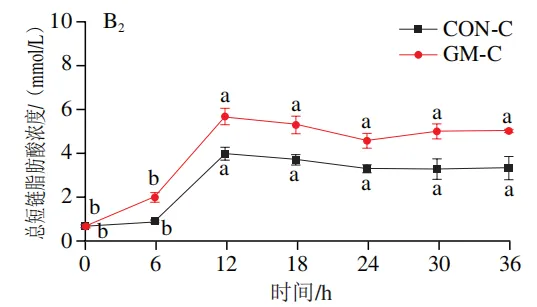
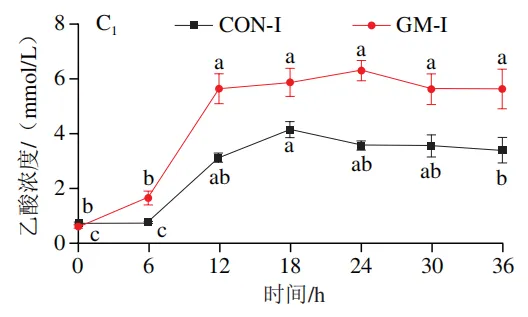
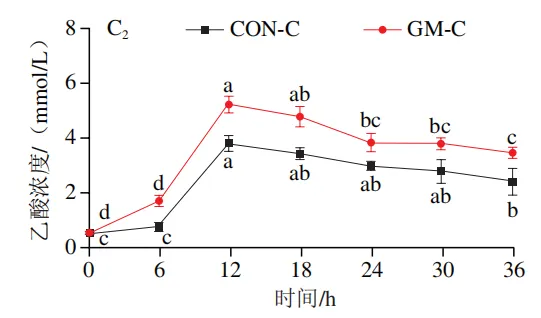
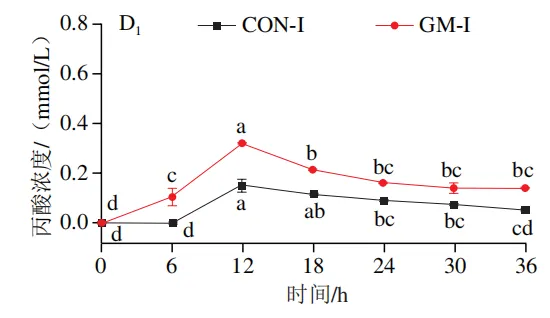
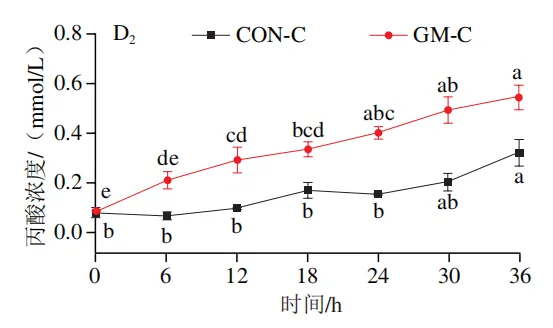
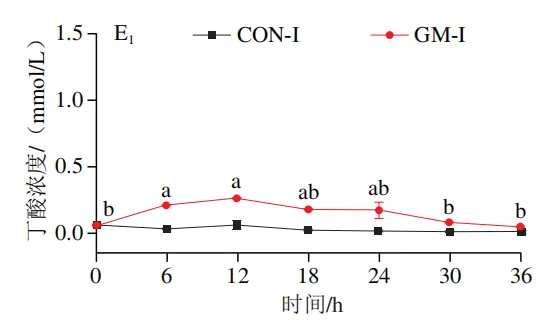
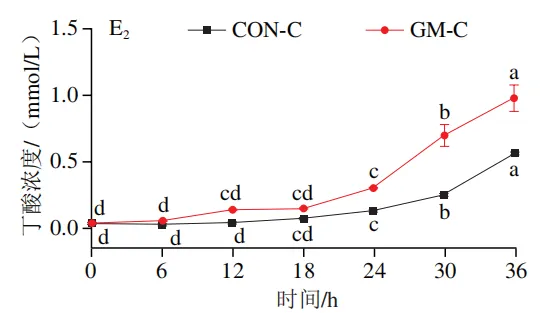
如图4A所示,4 个实验组发酵液中乳酸含量均在发酵6 h后显著降低(P<0.05)。在发酵6~36 h,回肠和结肠对照组发酵液中乳酸含量无显著变化,均保持较低的水平;回肠和结肠半乳甘露聚糖组乳酸含量在发酵12 h后显著上升(P<0.05),且在发酵12~24 h呈现较高水平,随后略有下降。如图4B所示,4 个实验组发酵液中总短链脂肪酸含量变化呈现相似趋势,即在发酵12 h后显著上升,在发酵12~36 h后保持较高水平。其中,回肠和结肠半乳甘露聚糖组发酵后各时间点总短链脂肪酸含量始终高于对照组。进一步分析各实验组乙酸、丙酸和丁酸含量,以明确各类短链脂肪酸的变化。在回肠发酵液中,对照组和半乳甘露聚糖组乙酸含量在发酵12 h后显著上升(P<0.05),在发酵12~36 h后无显著变化,且半乳甘露聚糖组乙酸含量始终高于对照组(图4C1)。回肠对照组和回肠半乳甘露聚糖组丙酸含量在发酵0~12 h逐渐增加,在发酵12 h时达到最高值,随后逐渐降低,且半乳甘露聚糖组丙酸含量始终高于回肠对照组(图4D1)。回肠对照组丁酸含量在整个发酵期无显著变化,而回肠半乳甘露聚糖组丁酸含量在发酵6 h后显著增加(P<0.05),随后逐渐下降至0 h水平(图4E1)。在结肠发酵液中,对照组和半乳甘露聚糖组的乙酸含量在发酵12 h后显著上升(P<0.05),随后逐渐下降,且结肠半乳甘露聚糖组的乙酸含量始终高于结肠对照组(图4C2)。结肠对照组和结肠半乳甘露聚糖组的丙酸和丁酸含量随发酵时间的延长而逐渐增加,且结肠半乳甘露聚糖组的丙酸和丁酸含量始终高于结肠对照组(图4D2、E2)。
 |  |  |
 |  |  |
 |  | |
 |  |

05
发酵液中微生物α多样性的变化
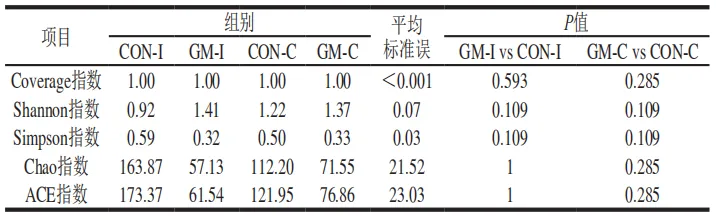
本研究通过Shannon指数和Simpson指数评估发酵液微生物的多样性,通过Chao指数和ACE指数评估发酵液微生物的丰度,发酵液中微生物α多样性的分析结果如表1所示。半乳甘露聚糖的添加使回肠和结肠发酵液中的微生物的Shannon指数略有增加,Simpson指数、Chao指数和ACE指数有所降低,但未达到统计学显著水平。
06
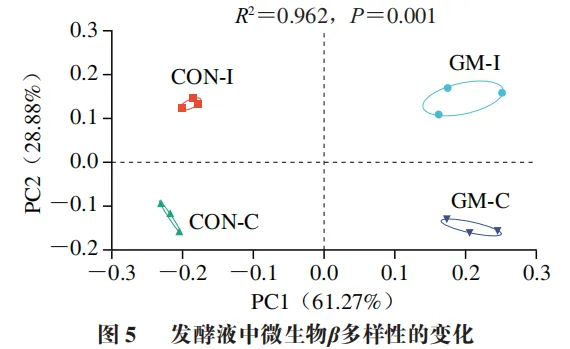
发酵液中微生物β多样性的变化
基于Bray-Curtis距离对回肠和结肠发酵液微生物测序结果进行主坐标分析(PCoA),发现4 个实验组的微生物结构存在明显分簇(图5)。ANOSIM分析同样显示,4 个实验组间存在显著差异(R2=0.962、P=0.001)。上述结果提示,猪回肠和结肠微生物结构有所不同,且半乳甘露聚糖的添加改变了猪回肠和结肠微生物的β多样性。
07
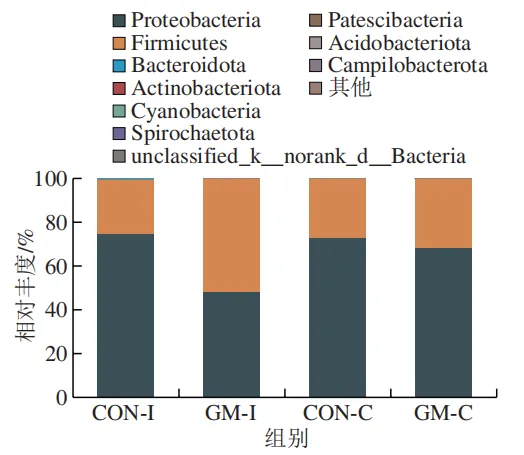
体外发酵液中微生物组成的变化
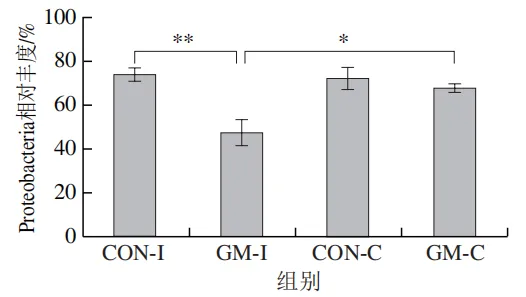
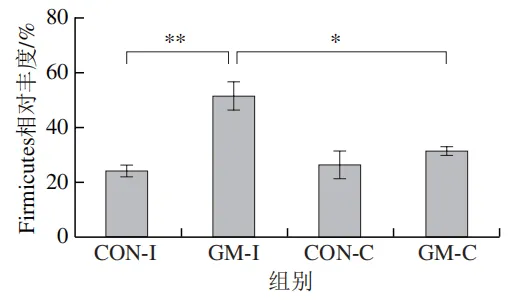
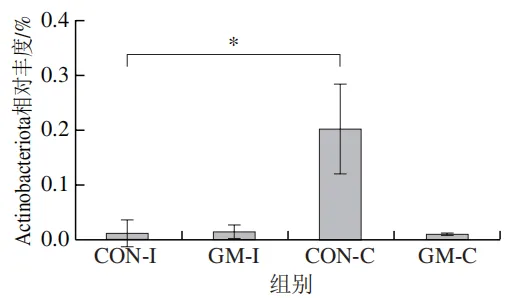
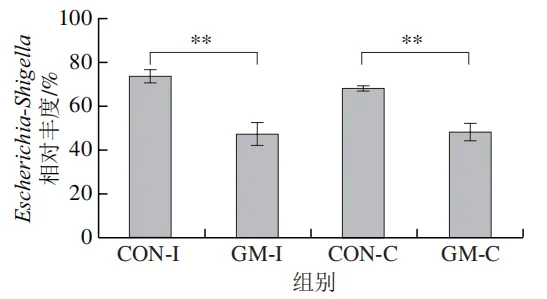
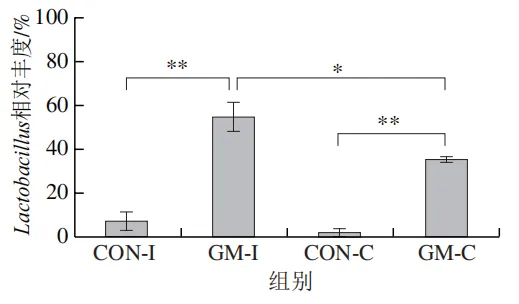
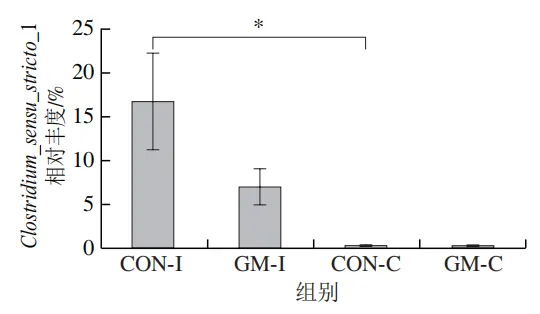
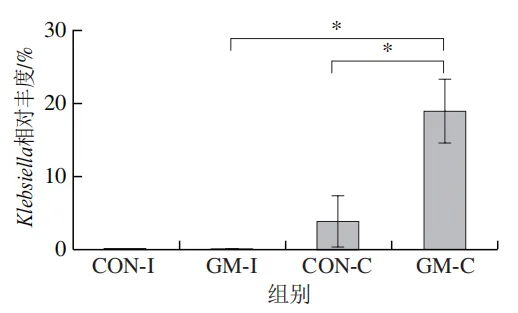
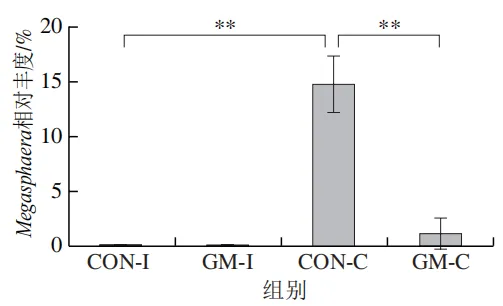
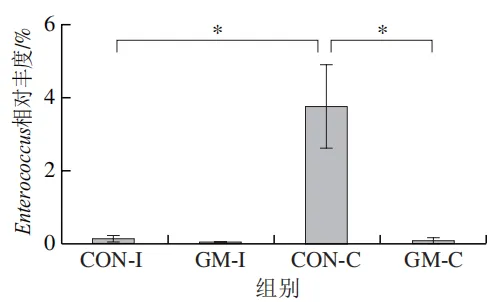
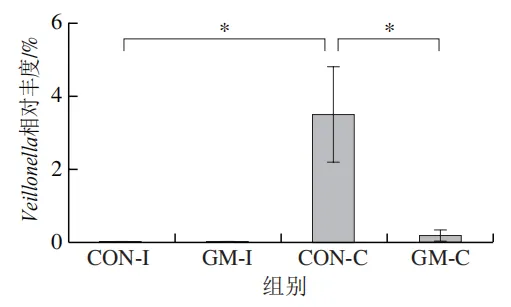
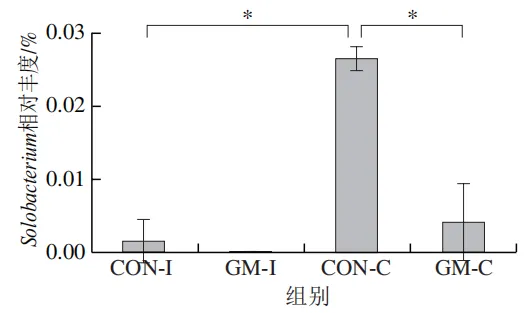
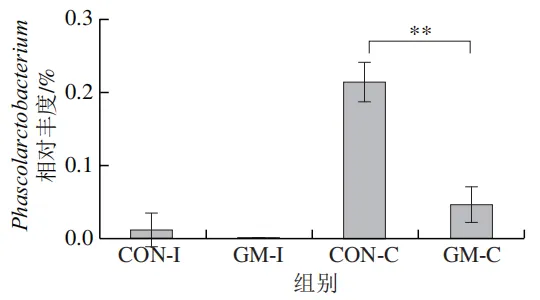
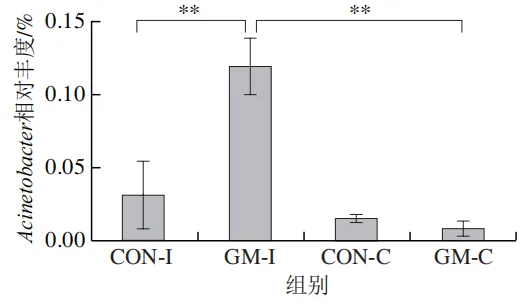
为了分析半乳甘露聚糖对猪回肠和结肠微生物组成的影响,本实验对发酵液中微生物的门水平和属水平的相对丰度进行统计分析。在门水平上,变形菌门(Proteobacteria)、厚壁菌门(Firmicutes)和拟杆菌门(Bacteroidota)为回肠发酵液的优势菌门,Proteobacteria、Firmicutes和放线菌门(Actinobacteriota)为结肠发酵液的优势菌门(图6)。与回肠对照组相比,结肠对照组Actinobacteriota的相对丰度较高(P<0.05)。在回肠发酵液中,半乳甘露聚糖的添加显著降低了Proteobacteria的相对丰度,增加了Firmicutes的相对丰度(P<0.05)。与回肠半乳甘露聚糖组相比,结肠半乳甘露聚糖组Proteobacteria的相对丰度较高(P<0.05),Firmicute的相对丰度较低(P<0.05)。
 |  |
 |  |
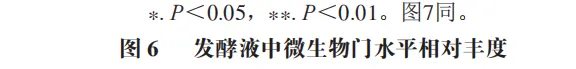
 |  |  |
 |  |  |
 |  |  |
 |  |
08
讨 论
8.1 回肠和结肠微生物对半乳甘露聚糖的利用差异
微生物在宿主肠道呈现区室化的空间分布,即不同肠段微生物的结构和代谢特征不同,进而对肠腔中碳水化合物的代谢表现出明显差异。为了明确回肠和结肠微生物对半乳甘露聚糖的利用差异,本实验研究发酵过程中发酵液pH值、半乳甘露聚糖、有机酸、氨氮以及菌体蛋白含量的变化规律。结果表明,尽管回肠、结肠半乳甘露聚糖组发酵液中半乳甘露聚糖的含量在发酵0~6 h后无显著变化,但其pH值显著下降,乙酸、丙酸含量有所上升,提示发酵前期回肠和结肠微生物主要利用发酵液中原有的营养物质进行代谢。此外,回肠和结肠发酵液中半乳甘露聚糖含量分别在发酵18 h和12 h后显著下降,且结肠发酵液中半乳甘露聚糖含量在发酵12~36 h始终低于回肠发酵液。上述结果表明,尽管两类微生物均可利用半乳甘露聚糖,但结肠微生物对半乳甘露聚糖的利用能力更强。造成上述现象的原因可能涉及两方面:1)与结肠微生物相比,回肠微生物的数量较少,且主要以蛋白发酵菌为主,这一生理特性可能直接影响了回肠微生物对半乳甘露聚糖的利用;2)厌氧发酵液中通常添加有酪蛋白胨等氮源,以确保微生物的生长繁殖,因此氮源的添加可能促进了回肠发酵液中蛋白分解菌的生长繁殖,使其在发酵初期生长较为活跃。此外,本研究发现两种发酵液中菌体蛋白含量均在6~12 h达到较高水平,该现象与同期pH值的显著下降及短链脂肪酸含量的显著上升趋势一致,提示半乳甘露聚糖的益生元作用在发酵6~12 h后较为强烈,有效促进了微生物增殖,进而驱动短链脂肪酸的合成并降低发酵液pH值。
8.2 半乳甘露聚糖对回肠和结肠有机酸合成的影响
肠道微生物能够帮助宿主将不能消化的碳水化合物代谢为乳酸和短链脂肪酸,这些有机酸既有助于调节肠道pH值,为乳酸菌、双歧杆菌等益生菌的生存提供适宜条件,又可为肠上皮细胞提供能量、调节肠道免疫功能。本实验发现,乳酸和乙酸是回肠和结肠微生物以半乳甘露聚糖为底物发酵的主要有机酸代谢产物。吴鹏晖等以猪小肠黏膜微生物为接种物,以低聚半乳糖或低聚甘露糖为唯一碳源的体外厌氧发酵实验同样观察到了类似的现象。上述结果提示,猪小肠和大肠微生物中均存在大量乳酸和乙酸产生菌。有趣的是,回肠和结肠半乳甘露聚糖组发酵液中乳酸含量均在发酵6 h后降至最低,在发酵12 h后显著上升。乳酸是一种中间代谢产物,既可被肠道微生物通过乳酸代谢途径转化为丙酮酸等物质,也可被肠道中多种互养菌转化为丙酸和丁酸。因此,在发酵初期,发酵液中的乳酸可能被优先转化为其他代谢物质,导致乳酸含量显著降低。而后,发酵液pH值的下降为乳酸菌的生长繁殖提供了适宜条件,进而促进了乳酸菌的增殖以及发酵液中乳酸的积累。
8.3 半乳甘露聚糖对回肠和结肠微生物多样性与组成的影响
为了评估半乳甘露聚糖对不同肠段微生物的调节作用,首先比较了两种发酵液的微生物α多样性的变化。α多样性体现了物种丰度与多样性,衡量指标包括Coverage指数、Shannon指数、Simpson指数、Chao指数和ACE指数。于颖等利用体外发酵技术探究了瓜尔豆胶半乳甘露聚糖对肥胖小鼠粪便微生物α多样性的影响,发现瓜尔豆胶半乳甘露聚糖显著降低了肥胖小鼠粪便微生物的Shannon指数、Simpson指数、Chao指数和ACE指数。此外,小鼠体内实验结果表明,具有益生元潜力的菊粉能够显著降低2型糖尿病小鼠肠道菌群的Shannon指数和Chao指数。本研究发现,添加半乳甘露聚糖使回肠和结肠发酵液的Simpson指数、Chao指数和ACE指数略有降低,但未达到统计学显著水平,这可能归因于发酵时长的差异。
09
结 论
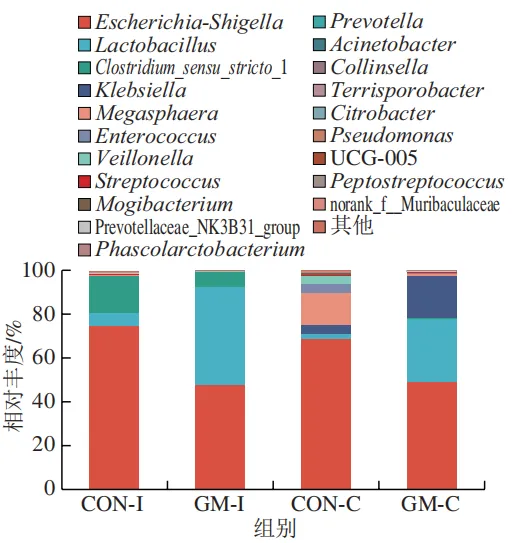
本研究借助体外发酵技术探究了不同肠段微生物对半乳甘露聚糖的利用效率和响应差异。结果发现,回肠和结肠微生物对半乳甘露聚糖的利用效率在发酵6~12 h较高,在此阶段pH值显著下降,短链脂肪酸、氨氮和菌体蛋白含量显著增加。其中乳酸和乙酸是回肠和结肠微生物利用半乳甘露聚糖进行厌氧发酵的主要有机酸代谢产物,丙酸和丁酸则是两类微生物在厌氧发酵过程中主要的差异有机酸代谢产物。半乳甘露聚糖改变了回肠和结肠发酵液微生物的多样性和组成,导致Lactobacillus相对丰度显著上调,Escherichia-Shigella相对丰度显著下调。上述研究表明,半乳甘露聚糖具有益生元潜力,能够优化肠道菌群结构、促进短链脂肪酸产生,且其对结肠微生物的调节作用更强。尽管如此,本实验仍存在一定局限性。例如,本研究采用的体外发酵模型虽可在一定程度模拟动物体内的厌氧环境,但忽略了肠道蠕动、宿主分泌物等在厌氧发酵过程中的作用,未来需在动物模型中进行进一步验证。此外,本研究发现回肠和结肠微生物对半乳甘露聚糖的利用差异主要体现在丙酸和丁酸代谢途径,未来可借助宏基因组技术深入解析造成丙酸、丁酸代谢差异的关键功能基因或酶学途径。
引文格式:
蒋雨欣, 张亭, 陈亚楠, 等. 体外法分析不同肠段微生物对半乳甘露聚糖的利用规律[J]. 食品科学, 2025, 46(21):126-136. DOI:10.7506/spkx1002-6630-20250611-072.
JIANG Yuxin, ZHANG Ting, CHEN Yanan, et al. In vitro analysis of the utilization patterns of galactomannan by microbiota from different intestinal segments[J]. Food Science, 2025, 46(21): 126-136. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-20250611-072.
点击下方阅读原文即可查看文章相关信息。
实习编辑:杨倩;责任编辑:张睿梅。点击下方阅读原文即可查看全文。图片来源于文章原文及摄图网
近期研究热点

随机文章
-
 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
- 【中绿农】《中国农业产业发展报告2025》:深入剖析中国农业产业韧性及竞争力
- 华南农业大学校长,去遗传发育所了
- 沈阳农业大学副教授第一作者在一区top期刊发表研究成果,取得塑料生物降解研究领域的新进展
- 爱尔兰农业、食品和海事部国务部长率领高级别贸易代表团访华,亮相青岛
- 未来农业政策,更愿意支持那些能'点亮一片'的项目,而不是'昙花一现'的个案
- 品牌农业社群营销:如何把1000个铁粉变成100个忠实客户?
- 氢农业特别适合于设施园艺农业
- 未来农业来了!这家上海公司在集装箱里种菜,用水不到传统农业5%
- 2024-2025农业年:安哥拉出口水果384吨
- 江南大学教授,入选农业农村部神农青年英才,发表Nature Communications揭示微生物的耐酸机制及代谢调控方面的研究成果
