3.1 剪草后日本结缕草的生理生化分析
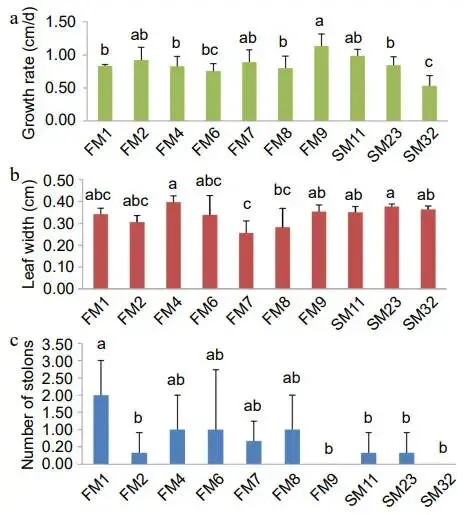
为筛选出再生速率差异最大的日本结缕草材料,对10份材料在修剪后进行了生长速率、叶宽及匍匐茎数量的测量。结果显示,FM9生长速率最快,SM23叶片最宽(0.38 cm),而FM1的匍匐茎数量最多(2条)。综合考虑,选择具有最多匍匐茎数量和相对较快生长速率(0.84 cm/d)的FM1作为高再生率材料,同时选择生长速率最慢(0.53 cm/d)且匍匐茎数量最少的SM232作为慢再生率材料用于后续实验 (图1)。
图1. 不同日本结缕草的生长情况。(a) 日本结缕草生长速率。(b) 日本结缕草叶宽。(c) 日本结缕草匍匐茎数量。误差棒代表±标准差。采用单因素方差分析进行显著性分析 (p < 0.05)。
3.2 日本结缕草修剪后再生时间点的形态学验证
再生启动: FM1的顶端分生组织在修剪后2小时出现并生长,而SM232则在12小时后才发生。
器官形成: FM1在修剪后6小时已出现叶原基和新分蘖,而SM232在24小时后才开始出现叶原基,48小时后出现新分蘖。
再生差异: FM1的叶原基和新分蘖数量均显著高于SM232,表明其新器官发生阶段更早,再生速度更快。
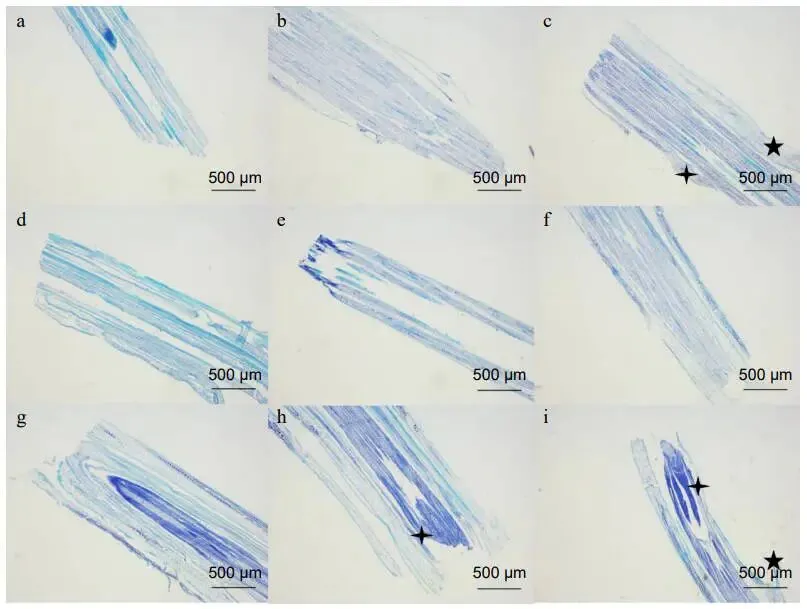
通过对FM1和SM232的生物切片进行不同时间的观察,验证了其再生时间点。FM1的顶端分生组织在修剪后2小时即出现并生长,并在6小时后同时观察到叶原基和新分蘖的形成。相比之下,SM232的顶端分生组织在12小时后才出现,叶原基在24小时后开始形成,新分蘖则在48小时后出现。结合形态学和微观结构变化推断,FM1的新器官发生阶段可能在修剪后6小时,而SM232则在24小时内。FM1的叶原基和新分蘖数量均显著高于SM232,证实了其更快的再生能力 (图2)。
图2. FM1和SM232修剪后顶端分生组织的微观结构变化。(a)−(c) FM1修剪后0、2和6小时的顶端分生组织。(d)−(i) SM232修剪后0、2、6、12、24和48小时的顶端分生组织。箭头指向叶原基;箭头指向分蘖。
3.3 日本结缕草再生过程的转录组学分析
表达模式: FM1的基因表达在修剪后2小时急剧上调,而SM232的基因表达变化在6至12小时后逐渐明显。
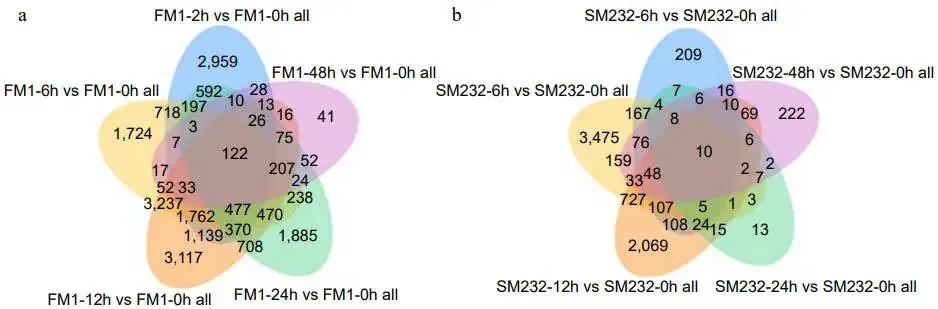
差异基因: FM1和SM232在不同时间点获得了大量差异表达基因(DEGs),其中FM1-12h和SM232-6h是差异基因数量变化最大的时期。
功能富集: FM1-2h的DEGs主要富集在初级代谢、激素信号转导和光合作用等通路。SM232-12h的DEGs则主要富集在核糖体、光合作用和翻译等过程。
响应延迟: 功能富集分析显示,FM1修剪后2小时的富集通路与SM232修剪后12小时的相似,表明SM232的响应存在延迟。
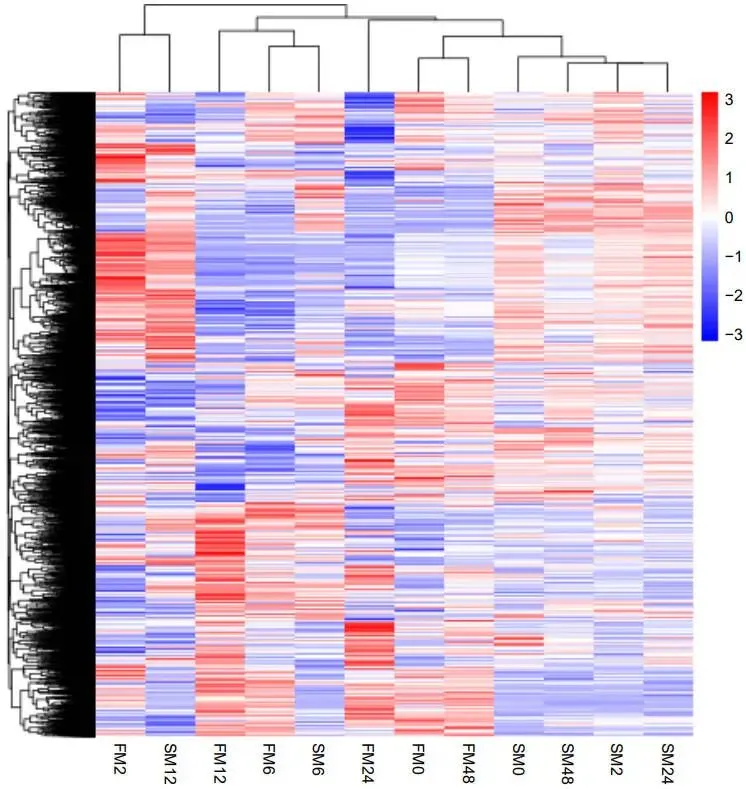
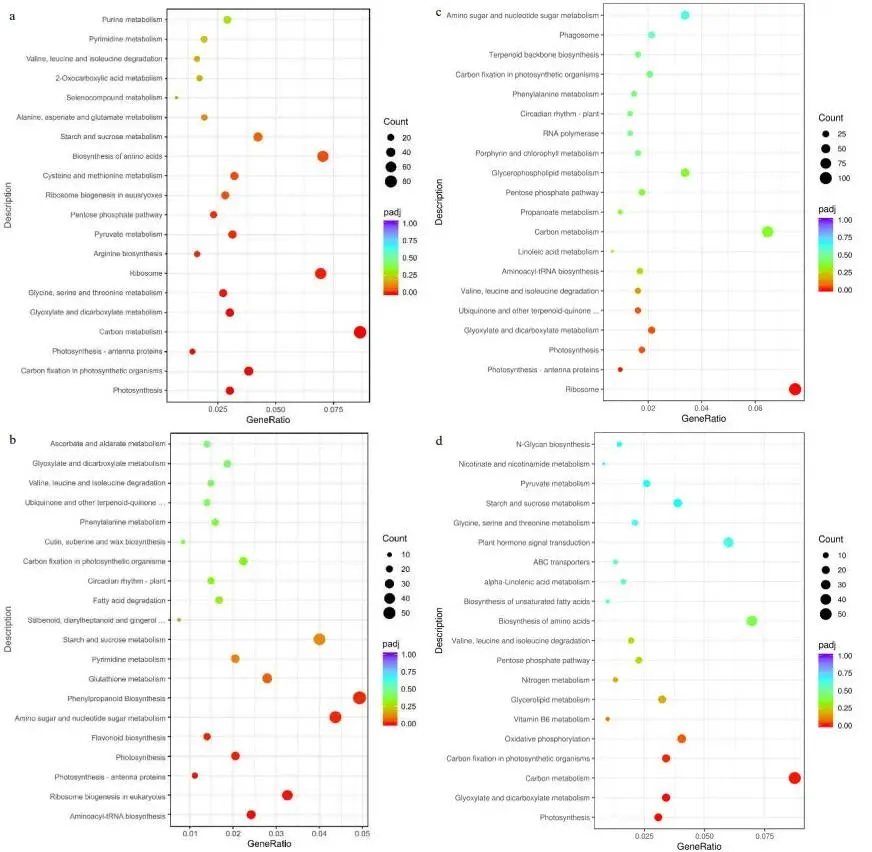
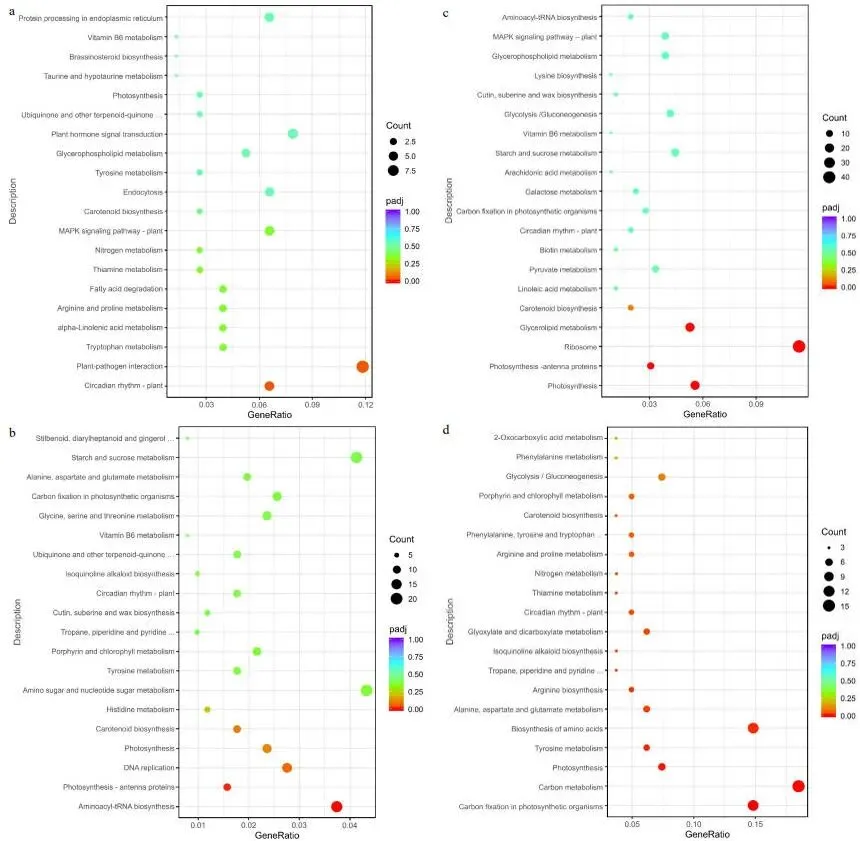
转录组分析显示,FM1的基因表达在修剪后2小时(FM1-2h)出现急剧上调,而SM232的基因表达变化趋势在6和12小时后(SM232-6h和SM232-12h)才逐渐显现 (图3)。差异表达基因(DEGs)的维恩图分析表明,两个材料在不同时间点均有大量DEGs产生 (图4a, b)。结合KEGG富集分析,差异基因数量变化最大的时期分别为FM1-12h和SM232-6h (图5, 6)。对DEGs的功能注释进一步揭示,FM1-2h的DEGs主要富集于碳代谢、激素信号转导、氨基酸生物合成及光合作用等通路。而SM232-12h的DEGs则高度富集于肽代谢、翻译、核糖体及光合作用相关GO类别。值得注意的是,GO和KEGG分析均显示,FM1修剪后2小时的富集通路与SM232修剪后12小时的相似,表明SM232的转录组响应存在显著延迟。
图3. 基于12个文库(FM1-2h, FM232-6h, SM232-2h等)配对比较的DEGs聚类分析。DEGs表达量的倍数变化进行层次聚类并以热图展示。表达水平的差异由色阶表示,从蓝色(下调表达)到红色(上调表达),如色阶条所示。由novomagic (https://magic.novogene.com) 制作。
图4. FM1和SM232修剪后不同时间差异基因的维恩图。(a) FM1修剪后2, 6, 12, 24, 48小时的差异基因数量。(b) SM232修剪后2, 6, 12, 24, 48小时的差异基因数量。
图5. FM1修剪后不同时间差异表达基因的KEGG富集气泡图。(a) FM1-2h与FM1-0h比较的DEGs的KEGG通路分析。(b) FM1-6h与FM1-0h比较的DEGs的KEGG通路分析。(c) FM1-12h与FM1-0h比较的DEGs的KEGG通路分析。(d) FM1-24h与FM1-0h比较的DEGs的KEGG通路分析。
图6. SM232建成后不同时间差异表达基因的KEGG富集气泡图。(a) SM232-2h与SM232-0h比较的DEGs的KEGG通路分析。(b) SM232-6h与SM232-0h比较的DEGs的KEGG通路分析。(c) SM232-12h与SM232-0h比较的DEGs的KEGG通路分析。(d) SM232-48h与SM232-0h比较的DEGs的KEGG通路分析。
3.4 再生过程中的激素变化
激素含量: FM1中的IAA含量显著高于SM232。两种材料的IAA含量在修剪后2小时下降,6小时后回升。CTK含量无显著差异。
基因表达: FM1中IAA和CTK相关基因均发生变化,而SM232中仅IAA相关基因有变化。
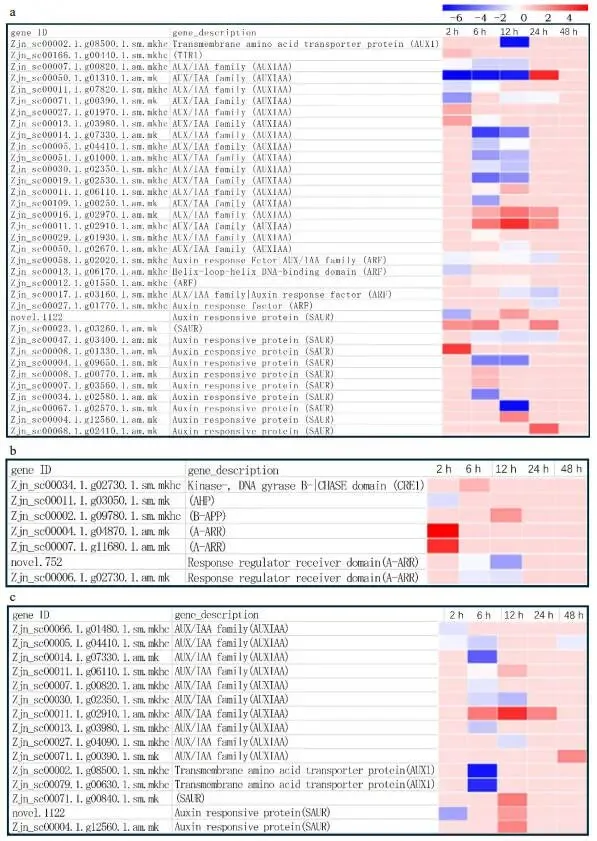
核心基因:AUX/IAA家族基因是变化最显著的,在FM1中鉴定到17个,SM232中10个,多数在6h和12h下调。CTK相关基因的表达变化仅在FM1中检测到。
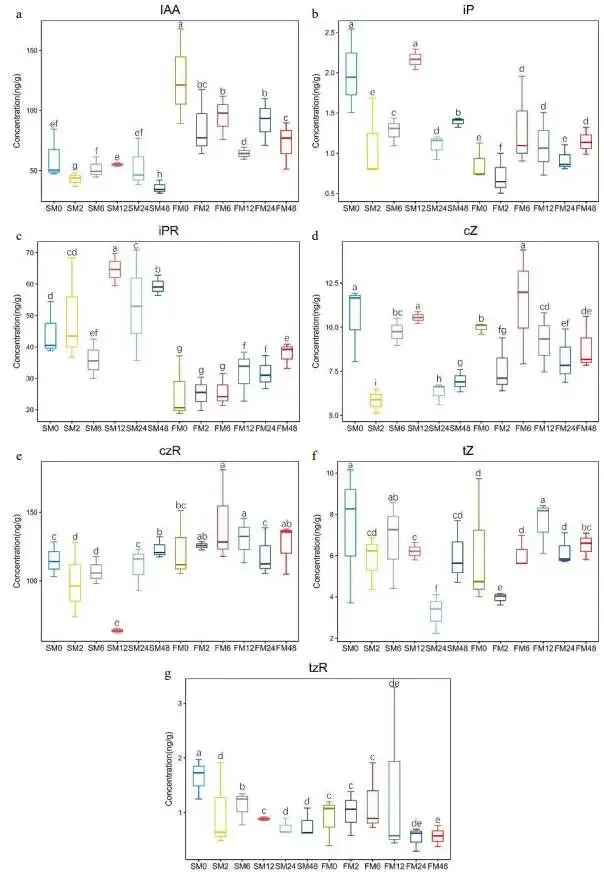
对两种材料中激素(IAA和CTK)含量的测定结果显示,FM1中的IAA含量显著高于SM232。两种材料的IAA含量均在修剪后2小时下降,6小时后再次升高。对于SM232,IAA含量在12小时达到峰值后逐渐下降;而FM1的IAA含量则在12小时降至最低后回升。两种材料间的CTK含量无显著差异,但SM232中的iP和iPR含量高于FM1 (图7)。在基因表达层面,FM1中IAA和CTK相关基因的表达均发生变化,而在SM232中仅发现生长素相关基因的表达变化,未检测到CTK相关基因的变化。在两种材料中,表达变化最显著的是AUX/IAA家族基因,FM1中鉴定到17个,SM232中鉴定到10个。这些基因的表达多数在修剪后6小时和12小时被下调。此外,在两种材料中也鉴定到SAUR家族基因(FM1中11个,SM232中3个),其表达变化主要集中在6小时或12小时。AUX1在两种材料中均呈显著下调趋势。FM1中还鉴定到5个ARF家族基因,在修剪后的不同阶段均处于下调状态 (图8a, c)。与IAA相关基因不同,CTK相关基因的表达变化仅在FM1中被检测到,其变化在修剪后短时间(2小时)内更为明显,有2个基因显著上调,1个基因下调。此外,另有4个基因在6小时和12小时分别上调或下调,但在长时间(24和48小时)内,所有这些基因的表达均未发生变化 (图8b)。
图7. FM1和SM232修剪后IAA和CTK的变化趋势。(a) FM1和SM232修剪后的IAA含量。FM1和SM232修剪后的CTK含量。(b) iP, (c) iPR, (d) cZ, (e) czR, (f) tZ, (g) tzR。
图8. FM1和SM232中IAA和CTK相关激素的基因表达。(a) FM1修剪后不同时间IAA相关基因表达的变化。(b) FM1修剪后不同时间CTK相关基因表达的变化。(c) SM232修剪后不同时间IAA相关基因表达的变化。
独特的研究视角与模型: 区别于以往多数集中于植物组织培养(in vitro)的研究,本研究首次聚焦于日本结缕草(Zoysia japonica)在模拟实际养护情景的机械损伤(修剪)后,其器官再生(in vivo)的分子机制。这使得研究结果更贴近实际应用,具有直接的指导意义。
整合多组学的系统性分析: 本研究首次将时间序列的转录组学分析与靶向激素代谢组学分析相结合,系统地描绘了日本结缕草修剪后再生过程中的动态分子调控网络。这种整合分析方法能够更全面地揭示从基因转录到激素信号响应的完整调控链条。
创新的比较研究设计: 研究巧妙地选取了两个具有显著再生速率差异的基因型(快速再生的FM1与慢速再生的SM232)进行对比分析。这种设计有效地将分子层面的差异(如特定基因的表达、激素含量的变化)与宏观的再生表型直接关联,为筛选关键调控因子提供了强有力的证据。
关键调控通路的新发现: 本研究一个重要的创新发现是揭示了细胞分裂素(CTK)信号通路在快速再生过程中的关键作用。研究发现,CTK相关基因的表达变化仅在快速再生的FM1中被检测到,而在慢速再生的SM232中则没有,这表明CTK信号通路的激活可能是区分两种基因型再生能力差异的核心分子事件之一。
鉴定出新的候选基因资源: 研究成功鉴定出一批与日本结缕草再生能力密切相关的新候选基因,特别是LOB、AUX/IAA及SAUR等家族中的特定成员。这些新发现的基因不仅丰富了对植物再生机制的理解,也为未来通过分子育种技术改良草坪草的恢复能力提供了宝贵的基因资源。
- 日本结缕草是一种重要的暖季型草坪草,但其慢速再生速度限制了其应用。研究选取了再生速度差异显著的两个基因型FM1和SM232进行分析。
- 转录组分析表明,剪草后2小时和12小时是狗牙根响应剪草的关键阶段。FM1的再生相关基因显著上调,而SM232的蛋白相关基因持续下调。
- IAA(生长素)含量在FM1中显著高于SM232,且FM1的CTK(细胞分裂素)相关基因发生显著变化,而SM232未检测到CTK相关基因的变化。
- FM1的再生速度快于SM232,剪草后6小时内FM1开始形成叶原基和新分蘖,而SM232的再生过程延迟至24小时后。
- LOB家族蛋白相关基因在FM1中显著上调,表明其可能与狗牙根的侧生器官形成和再生能力密切相关。
- AUX/IAA家族基因在FM1中的表达显著高于SM232,且与狗牙根的再生速度密切相关。
- SAUR家族基因在FM1中表达较多,而SM232中仅有少量基因表达,这些基因可能参与调节细胞扩展及再生过程。
- ARF家族基因在不同阶段表达下调,与植物生长和发育中的生长素信号传导相关。
- 剪草后,FM1在短时间内表现出快速的生理反应和激素变化,而SM232的反应较慢,表明基因型差异显著影响再生机制。