重磅!中国农业大学发表《Cell》
- 2026-07-05 20:45:16

受精后卵裂形成桑葚胚,进一步发育为囊胚;囊胚内细胞团分化、重排,分化出内胚层与外胚层,两层紧贴形成二胚层胚盘,位于羊膜腔与卵黄囊之间,是未来胚胎本体的原始结构。胚盘形成的核心关键作用包含:1. 继续建立二胚层结构,奠定三胚层分化的基础;2. 构建胚胎微环境;3. 建立胚胎前后、背腹初始极性。
人类胚盘发育于胚胎第 3 周,此后原条出现,原肠运动正式启动。然而,鉴于该时期的特殊性,对于胚盘形成,原条发生的诱导信号、空间组织规律以及胚外组织在这些过程中对胚胎的命运调控到底是什么?相关问题长期未被解析。
2023年,中国农业大学生物学院魏育蕾团队与德克萨斯大学西南医学中心吴军团队、昆明理工大学谭韬团队通过建立干细胞模型,基于对早期胚胎发育囊胚形成时期Hypoblast与Epiblast的谱系互作关系,解析着床前胚胎发育的多谱系互作,为研究胚外胚内互作开辟了新型研究范式,该文章发表于2023年的Cell杂志“Dissecting embryonic and extraembryonic lineage crosstalk with stem cell co-culture”。

2026年6月24日,基于胚外谱系与胚胎互作研究系统,中国农业大学生物学院魏育蕾教授团队与中国科学院动物研究所团队合作,再次在Cell杂志上发表了题为“Reconstituting Human Primitive Streak Formation through Extra-Embryonic Cell Coordination”的研究论文。该研究基于2023年建立的多谱系互作研究范式,将发育时间进一步推后,引入了原肠运动时期胚外谱系,系统解析了羊膜样细胞、滋养层干细胞和胚外中胚层干细胞在原条样结构形成过程中的多谱系互作与调控,并进一步结合micropattern技术构建disc-Gastruloid(胚盘样类原肠模型),在体外重现原条样结构的有序发生过程。为研究胚胎植入后胚盘形成和胚外—胚内细胞互作提供了新实验平台。

生命早期发育并非由胚胎细胞独立完成,胚外组织不可或缺。胚胎植入前后,滋养层、羊膜、胚外中胚层等胚外组织相继出现,并与胚内上胚层细胞共同构成植入后早期胚胎结构。越来越多证据提示,胚外组织不仅提供结构和营养支持,还可能通过细胞间信号交流参与调控胚胎细胞命运决定与空间组织建立。然而,现有整合型干细胞胚胎模型多依赖细胞自发聚集和分化,谱系混杂,难以精确界定并解析多种细胞在原条形成中的特异性作用。为此,研究团队首次纯化原肠时期三类胚外谱系细胞,并系统评估不同胚外谱系对原条发生的作用。
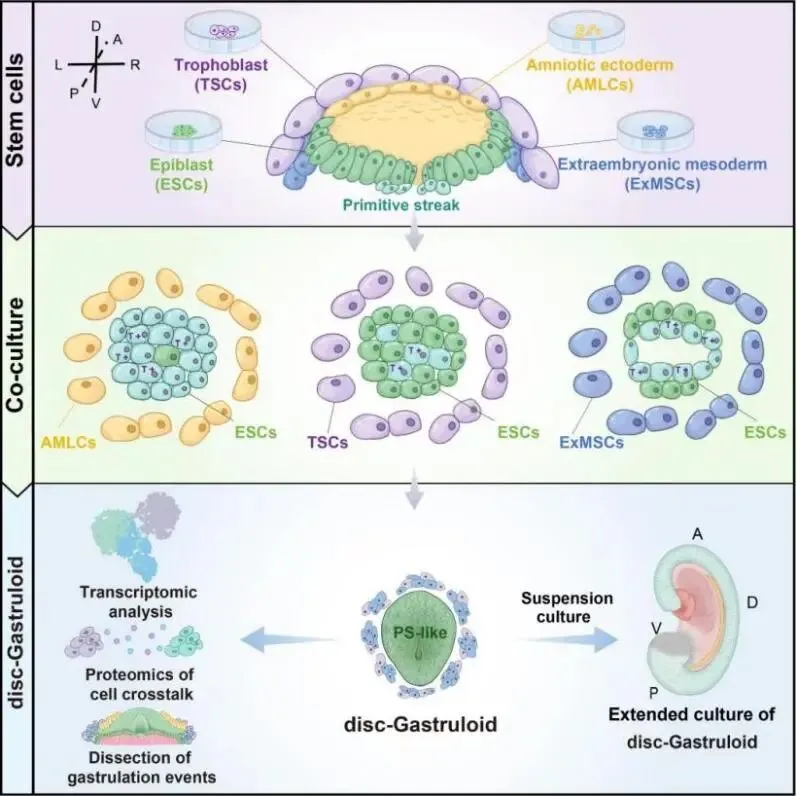
图1:Disc-Gastruloid 模型示意图:重构胚内/胚外空间互作,模拟原条样结构形成
一、关键发现:三类胚外细胞时序/空间协同调控原肠运动
TBXT 编码 T/Brachyury 转录因子,是原条、原结、脊索、早期中胚层特异性核心标志物,其时空表达严格同步于原条起始、延伸、细胞内迁全过程,二者协同调控原肠运动与体轴建成。三类胚外细胞(AMLCs、TSCs、ExMSCs)从分化启动、分化抑制、迁移定向三个独立环节分工调控 T⁺原条样细胞的产生与行为,共同协调着床后原肠期胚胎微环境调控网络。
1.1 类羊膜细胞 AMLCs:原条分化的起始诱导者(上游启动环节)
类羊膜细胞(AMLCs):能够诱导胚胎干细胞(ESCs) 产生原肠运动特征T+ 细胞。AMLCs 共培养条件下 ESCs 中出现明显 T+ 细胞群,并伴随TBXT、MIXL1等原肠运动相关基因表达。
1.2 滋养层干细胞 TSCs:原条分化的强抑制屏障(负向稳态调控)
滋养层干细胞(TSCs):对T+细胞的产生表现出抑制作用。即使在启动原肠发生的条件下,比如加入 WNT 通路激活剂 CHIR99021(CHIR),仍呈T+ 显著抑制现象,并促使更多细胞维持未分化状态。
1.3 胚外中胚层细胞 ExMCs:T⁺原条细胞的迁移导向者(下游运动调控)
胚外中胚层细胞(ExMCs):本研究发现,ExMSCs再胚胎发育过程中主要起迁移引导作用。研究团队基于2024年发表的“3D胚胎”系列论文(Cell,2024;Nature Cell Biology,2024;Cell Stem Cell,2025),解析体内胚外中胚层分子特征,优化ExMSCs新型稳定培养体系,通过优化基底、调节Activin A 和 FGFR1 抑制剂 PD173074 等条件,首次建立了可长期稳定培养的胚外中胚层干细胞( ExMSCs)。功能实验表明,ExMSCs 本身并不直接诱导 ESCs 表达 T,却能定向吸引已进入 T 阳性及上皮—间充质转化(EMT)状态的原条样细胞迁移。
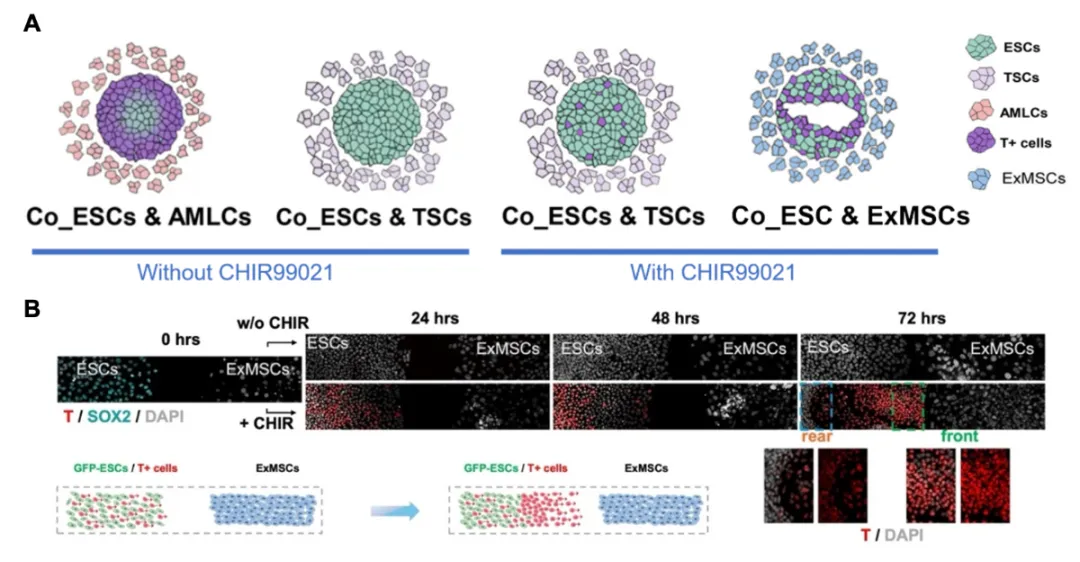
图2:胚外细胞协同调控 ESCs 原条样分化与 T⁺ 细胞迁移
二、 胚盘”重建:体外再现原条时空发生
在胚胎发育过程中,细胞命运的建立不仅依赖特定分子信号的精确调控,也高度受到空间位置和组织构型的影响。着床后胚胎中,胚内上胚层与羊膜、滋养层及胚外中胚层等胚外谱系共同构成复杂的时空微环境,为原条形成和原肠运动启动提供关键调控基础。
真实胚胎中,细胞命运不仅受分子信号调控,也高度依赖空间位置。基于上述关键发现中,对胚外谱系的重新构建与理解,研究团队结合micropattern 微图案化技术、Transwell 共培养体系和生物工程策略,重构原条形成时期胚内—胚外空间关系。通过模拟胚盘物理生理状态,最终构建出能够模拟着床后胚胎空间组织特征、并包含 AMLCs、TSCs 与 ExMSCs的体外胚盘模型。研究团队将该模型命名为Disc-Gastruloid。72 小时的共培养后,disc-Gastruloid 的胚盘样结构中逐渐出现沟槽状的类原条样区域。该区域细胞间连接减弱,并特异性表达 T/Brachyury、MIXL1 等原条及原肠运动相关标志物,提示模型中发生了类似天然胚胎原条形成过程的细胞迁移与命运转变。该模型能够动态再现原条形成过程中 EMT 启动、细胞迁移和谱系分化等关键事件。
Disc-Gastruloid 模型通过重构胚外细胞的空间与分子协同作用,在体外模拟了原条形成的关键过程,为研究早期胚胎发育生理、物理环境提供了全新的实验平台。
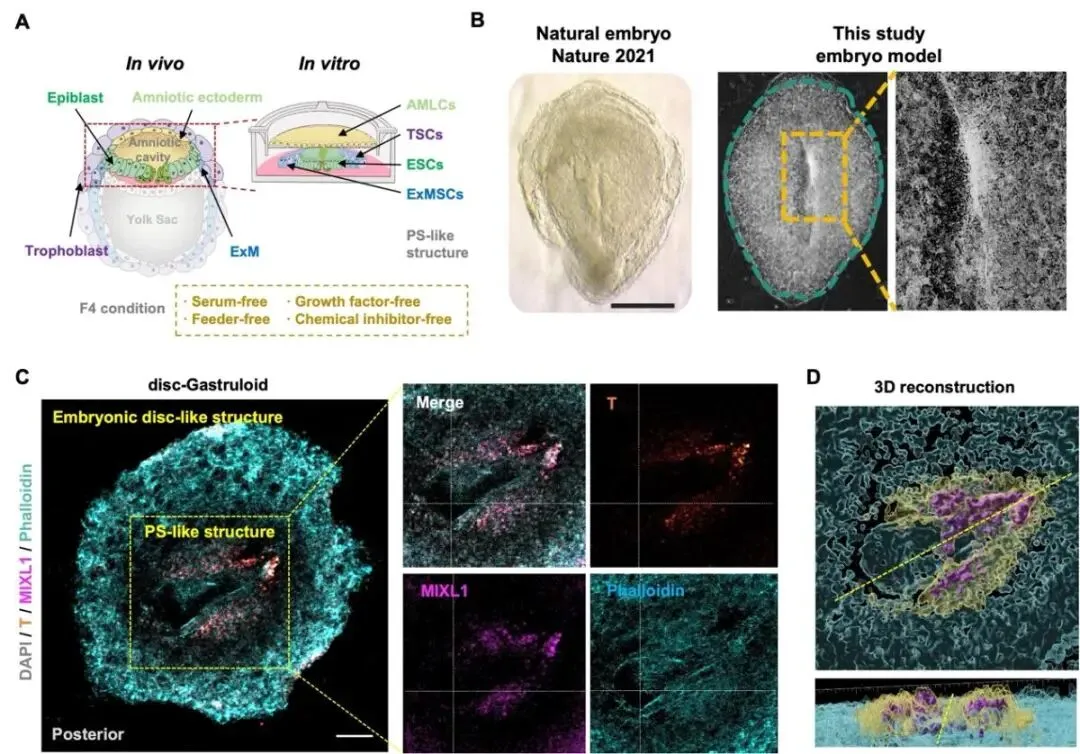
图3:体外空间重构胚盘样结构并诱导类原条样形态发生
三、胚盘发育:头尾背腹再现,神经管再生
为了进一步探索了Disc-Gastruloid 是否具备持续发育潜能,将胚盘样结构分离并进行低吸附悬浮延长培养,研究发现,超过 80% 的 disc-Gastruloid 能够在第 4 天自发形成伸长、不对称、弯曲且多层化的三维胚胎样结构。该结构中可观察到 SOX2⁺ 神经样细胞、SOX17⁺ 内胚层样管状结构,以及共表达 SOX2 和 T 的神经中胚层祖细胞样区域,并呈现类似早期胚胎前后轴和背腹轴的空间分布特征。部分结构前端还出现共表达 ISL1 和 cTnT 的心肌祖细胞样区域,并表现出自主节律性收缩。单细胞转录组分析显示,延长培养后的 Disc-Gastruloid 包含 14 类细胞群,其主要细胞类型与人CS9阶段(约受精后21天)胚胎中的对应细胞具有较高相似性。
研究结果表明,Disc-Gastruloid 虽仍与真正胚胎存在一定差异和异常,但能够在体外进一步发育为高度类似早期胚胎的复杂结构,为研究原肠运动发育机制提供了重要平台。
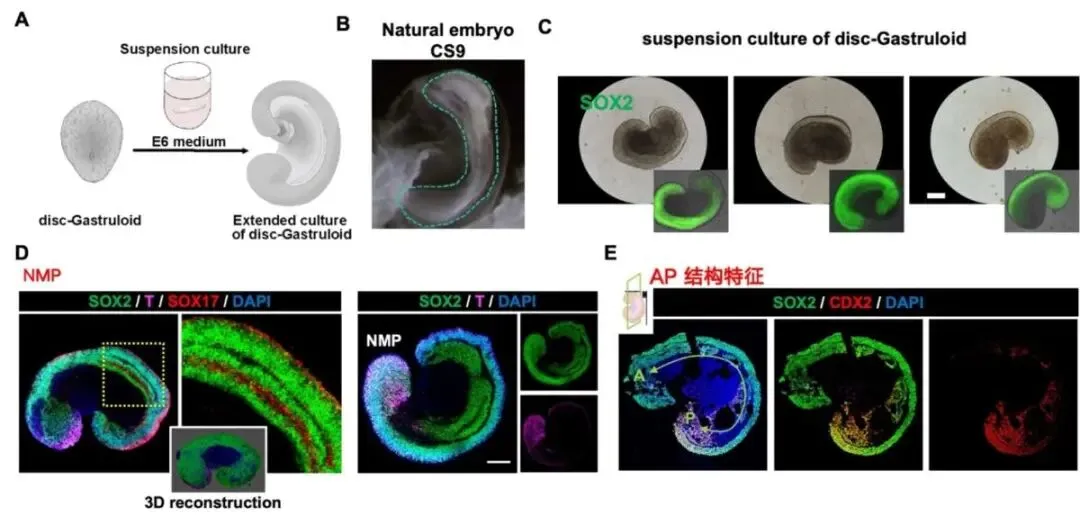
图4:延长培养的 disc-Gastruloid 形成类 CS9–CS10 胚胎样轴向结构
综上,本文解析了胚外组织对胚胎空间组织和命运决定的调控规律,系统揭示了三类胚外细胞在原条样结构形成中的分工与协同机制,充分阐明了早期发育的胚外组织在空间定位上的潜在生理/病理学意义。本研究继2023年魏育蕾团队在Cell上解析的“着床前囊胚时期,原始内胚层限制胎儿无序成长,规范胎儿正常增值分化”之后,进一步明确“着床后原肠胚时期,胚外组织对胎儿着床后的胚层分化、胚盘建立、原条形成有重要影响”。不同胚外组织缺失最终可致使胎儿早期流产、躯干缺损、神经管畸形、泌尿生殖系统发育畸形等现象。
该理论对农业动物胚胎发育效率、动物胚胎体外发育及移植、异种嵌合、多物种干细胞研究提供新的理论参考,也为未来利用家畜大动物构建复杂发育模型奠定基础。
本研究第一单位为中国农业大学生物学院、畜禽生物育种全国重点实验室、分子设计育种前沿科学中心、模式动物表型与遗传研究国家重大科技基础设施。中国农业大学生物学院的博士生张鑫、博士后陈乃馨参与到了项目中,与其他成员共同为该论文的共同第一作者。魏育蕾教授获得了教育部U40项目支持,国家重点研发计划、国家自然科学基金、动物生物技术育种全国重点实验室创新项目、中国农业大学基本科研业务费等项目资助。感谢模式动物表型与遗传研究国家重大科技基础设施(猪设施)提供支持,并感谢实验室全体成员的讨论。
招聘信息
中国农业大学魏育蕾课题组诚招博士、博士后(2-3名)
研究方向:
课题组围绕三个紧密关联的方向开展前沿探索:
方向一:人工胚胎--通过干细胞构建人工胚胎模型,利用多种工具系统解析胚胎发育分子机制。
方向二:生育繁殖--通过干细胞模型等解析人类/动物生殖发育基础,解析繁殖异常。
方向三:异种器官再造—通过异种嵌合等手段,解析人猪发育不平衡,克服人干细胞在猪体内发育障碍等分子机制,最终实现人类功能器官体外构建,用于再生医学。
本组郭文博老师(自动化系背景)招聘生物信息学与人工智能方向相关专业博士与博士后。
课题组老师信息网站
魏育蕾:https://cbs.cau.edu.cn/art/2022/11/1/art_31829_887741.html
郭文博:https://cbs.cau.edu.cn/art/2026/2/5/art_31830_1098647.html





