《食品科学》:中国农业科学院郑明明研究员、张逸副研究员等:神经酸生物合成、纯化及营养应用研究进展
- 2026-06-29 14:28:08

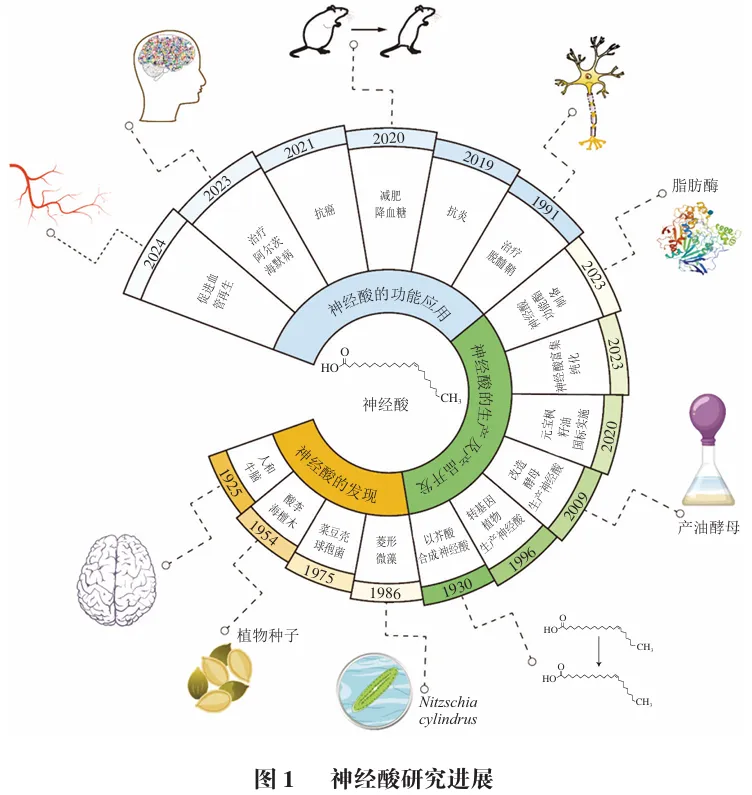
神经酸是一种长链单不饱和脂肪酸(顺式-15-二十四碳烯酸,ω-9),因其与神经系统密切相关而得名。神经酸可与鞘磷脂结合生成神经酰鞘磷脂,后者作为神经纤维的核心组成部分,是大脑白质及髓鞘的主要成分,在脊椎动物神经组织中发挥关键功能。此外,作为母乳中的天然成分之一,神经酸可促进神经细胞的增殖和分化,帮助婴儿神经系统的发育和成熟。如图1所示,神经酸及其衍生物具有细胞膜形成、信号传导、抗炎、调节血糖、促进髓鞘和血管再生等多种生理功能,其在食品、医药及保健品等领域具有广阔的发展与应用前景。
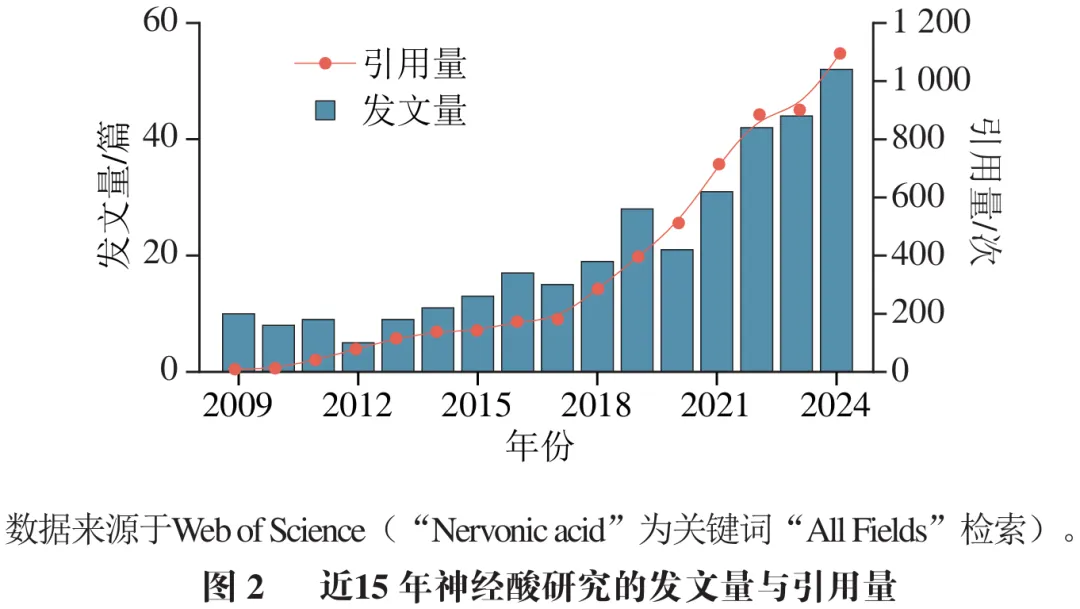
神经酸最初来源于牛和人类大脑中分离的不饱和脂肪酸,后又从鲨鱼油中提取制备,因此神经酸也被称为鲨鱼酸。然而,捕获鲨鱼制取神经酸会直接威胁鲨鱼种群的生存,破坏海洋生态系统的平衡,受海洋生态保护、公共卫生与安全、伦理道德等多方面因素制约,从鲨鱼中制取神经酸的方法已逐渐被放弃。1930年,Hale等提出以芥酸甲酯为原料,经还原生成芥酸醇、溴代、丙二酸乙酯化及水解脱羧等多步化学反应,合成了少量顺式结构的神经酸。1954年,Bounds等进一步提出以油酸为原料,经臭氧化、氧化、辛二酸甲酯缩合、碳链延长、还原及水解脱羧等步骤合成神经酸。这些早期尝试虽然为神经酸的化学合成奠定了研究基础,但仍存在工艺复杂、产率偏低、副产物多等产业难题。与此同时,1954年,Ligthelm等从南非的3种海檀木属植物中发现了神经酸,但植物提取法仍受限于植物资源分布不均、原料收集困难、成本高昂等瓶颈。1986年,Nichols等在圆柱形伪菱形藻(Nitzschia cylindrus)中也发现了神经酸,微藻较植物具有生长速度快、生长周期短的优势,但天然藻类的产油率偏低,且油脂中神经酸占比通常不足10.0%,仍难以满足工业生产需求。另一方面,James等报道了拟南芥中脂肪酸延长酶基因(FAEI)参与超长链脂肪酸的合成。Lassner等在植物霍霍巴中发现了与FAE1同源的β-酮酰基辅酶A合成酶(KCS)基因并导入欧洲油菜和拟南芥中,获得了油脂组成中神经酸质量分数分别为7.8%和1.6%的油菜籽和拟南芥籽。近年来,随着微生物合成和生物技术革新,通过基因工程、代谢工程及多组学整合等手段生物合成神经酸已逐渐成为研究热点。如图2所示,神经酸的年发文量从2009年的10篇逐年增加至2024年的52篇,引用频次也从2009年的9次增长到2024年的1 095次。因此,湖北工业大学生命科学与健康工程学院的高云旗,中国农业科学院油料作物研究所的张逸*和郑明明*等人对近15年的神经酸相关文献进行系统梳理和总结,探讨神经酸天然来源、生物合成途径、富集精制、功效评价和应用的研究进展,以期为神经酸的生物制造和产业化应用提供科学参考。
1
神经酸来源
1.1 植物来源
天然神经酸已在十字花科、铁青树科及无患子科等54个科136个属约200种植物中被发现,其中大多数植物神经酸含量不足其脂质组成的2.0%。基于神经酸的工业化生产潜力,表1总结了含油量超10.0%的7种木本植物和4种草本植物。其中,蒜头果产油率为58.0%~67.8%,油脂脂肪酸组成中神经酸占比高达67%,但蒜头果为我国二级保护植物,仅分布在云南和广西少部分区域。相比之下,元宝枫、文冠果和三花槭等槭属植物中神经酸在其脂质组成占比虽不足8.0%,但其分布范围广、种植量大、产油率高,仍被认为是神经酸的重要来源。元宝枫作为神经酸研究的热点植物之一,其种子脂肪酸积累及神经酸生物转化机制已通过转录组测序和核糖体足迹分析等手段逐渐被探明。

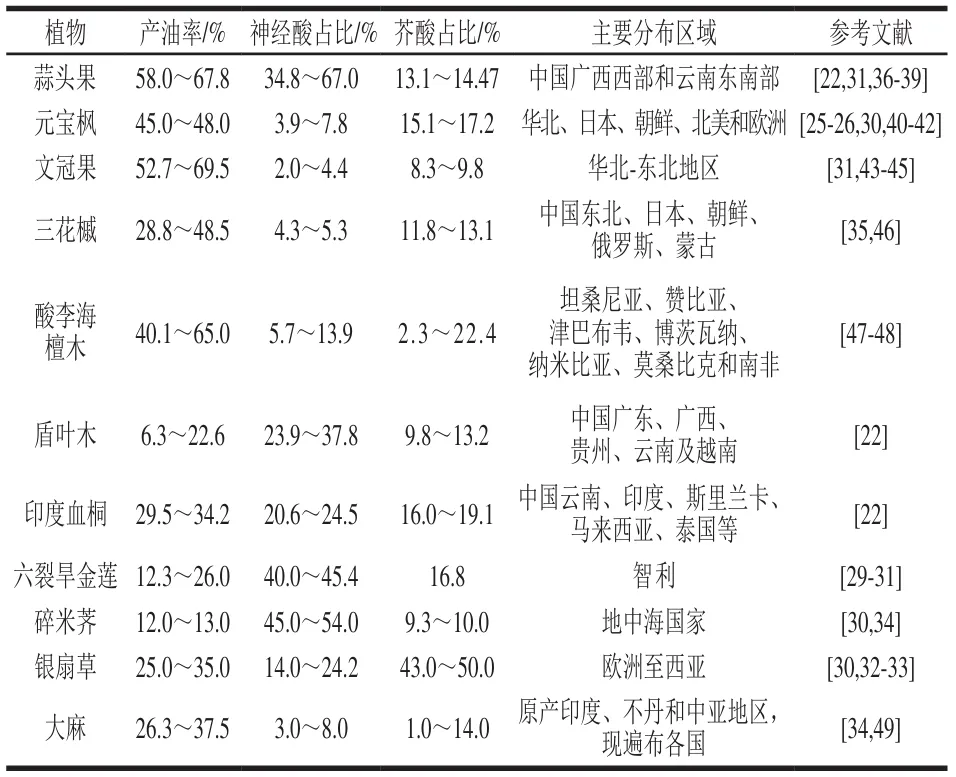
与木本植物相比,草本植物的种仁产油率更低,且存在产量波动大、收集困难和分布区域狭窄等问题。例如,六裂旱金莲种子产油率为12.3%~26.0%,神经酸占总脂肪酸的40.0%~45.4%,但其种子不易收集和播种。银扇草籽产油率为25.0%~35.0%,神经酸占比为14.0%~24.2%,被认为是具有神经酸产业化生产潜力的作物之一。然而,银扇草产量差异大(800~2 000 kg/ha),且银扇草籽油中芥酸占总脂质组成的43.0%~50.0%,存在心血管系统损害、器官损伤和生殖功能异常等风险;并且芥酸与神经酸分子结构相似,进一步增加了神经酸分离纯化的成本。碎米荠籽油神经酸占比可达45.0%~54.0%,且芥酸含量低,但碎米荠植株高度仅8~30 cm,对土壤要求严格,且种子易碎裂,限制了其大规模利用。
总体而言,受植物生长周期长、种仁收集困难以及籽油中神经酸含量普遍较低等因素限制,植物源神经酸提取率低,分离纯化工艺复杂,成本昂贵,但从天然植物中提取神经酸仍具有重要研究价值。一方面,从已知植物中筛选高神经酸、低芥酸含量的特异品系,是提升植物源神经酸的重要途径。例如,Su Yajing等筛选出一株三花槭,其种籽油中神经酸占比4.5%,芥酸占比低于1.0%,拓展了三花槭生产神经酸的应用潜力。另一方面,这些植物作为神经酸的种质资源,其脂肪酸代谢途径解析和系统生物学通路优化将为神经酸生物合成提供宝贵参考。
1.2 微生物来源
与植物生产相比,微生物具有生长速度快、生长周期短、不占用农业用地等优势,因此微生物法制备神经酸也被广泛报道。如表2所示,细菌中土拉弗朗西斯菌(Francisella tularensis)是可引起人和动物兔热病(又称野兔热)的一种革兰氏阴性、专性胞内寄生的短小杆菌,其体内转化的神经酸达脂质组成的11.2%~19.3%,但该菌对人类和动物具有高度致病性。而真菌中来自反刍动物瘤胃的厌氧菌Neocallimastix sp.、Orpinomyces sp.和Piromonas communis的安全性更高,但神经酸产量小于5.0%,且生长过程依赖厌氧环境,也限制了其产业化应用。此外,真菌中高山被孢霉菌株Mortierella capitata RD000969能够合成多种长链单不饱和脂肪酸,总脂质占其干质量的9.3%,但神经酸占比较低,仅为总脂质的6.9%。而植物病原菌菜豆壳球孢菌(Macrophomina phaseolina)对培养基成分表现出明显的响应,当培养基碳氮比为10∶1时,其总脂质占菌丝干质量的13.1%,其中神经酸占总脂质的39.0%;而当碳氮比升至80∶1时,总脂质含量下降至4.0%,但神经酸在总脂质组成中占比增加至48.8%。此外,Chaudhuri等也发现,通过调节培养基成分,可提高长孢被孢霉菌(Mortierella elongata)SC-208神经酸产量。栅藻、小球藻等部分藻类也具有一定的神经酸合成能力,其中,小球藻Nannochloris sp.QUCCCM31脂质组成中神经酸占比可达10.0%,并能耐受45 ℃高温及35~100 g/kg宽盐度范围,具有一定的神经酸生产潜力。另一种具有神经酸生产潜力的Mychonastes afer HSO-3-1,其油脂占干质量的53.9%,其中神经酸占比为3.8%。通过结合高氮源及低氮高光照两步法优化培养,神经酸产量提高至总脂肪酸的8.5%。而Andruleviciute等则发现在培养基中添加甘油可显著提高小球藻(Chlorella sp.)的生物量,并且发现其在不添加甘油的培养基上几乎不生产神经酸,而在添加2 g/L甘油条件下培养相同时间,其脂质中神经酸占比增加至3.1%。

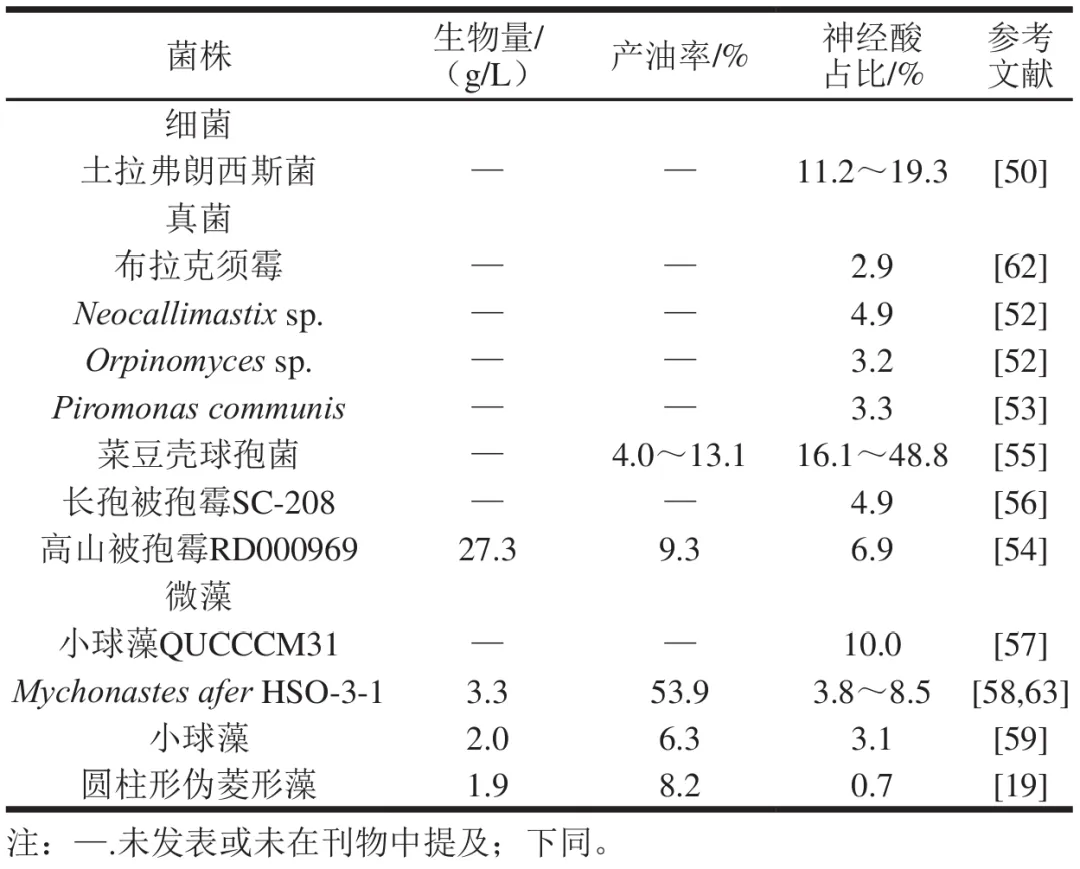
近年来,新的微生物如嗜热小球藻Chlorella thermophila MF179624和嗜酸乳杆菌Lactobacillus paragasseri MJM60645也被报道具有神经酸合成能力。总体而言,神经酸在微生物中广泛分布,但存在菌株安全性不明、培养条件要求高、油脂含量低等问题,后续可通过筛选安全的底盘菌株,结合代谢工程改造、优化菌株发酵工艺等方法提高神经酸产量。
2
神经酸的生物合成
2.1 生物合成机理
神经酸在动物、植物和微生物中的合成过程均可分为油酰辅酶A(C18:1,ω-9)的从头合成及其碳链延长两个阶段。如图3a所示,第1阶段中乙酰辅酶A在乙酰辅酶A羧化酶(ACCase)的作用下生成丙二酰辅酶A,并与酰基载体蛋白(ACP)结合,经脂肪酸合成(FAS)模块逐步延长碳链生成硬脂酰-ACP并去饱和化为油酰辅酶A。然而,不同生物体内硬脂酰-ACP去饱和为油酰辅酶A的过程存在显著差异,例如,植物中硬脂酰-ACP首先在去饱和酶的作用下转化为油酰-ACP,并水解生成游离脂肪酸,再通过酯化形成油酰辅酶A。在酵母中,硬脂酰-ACP首先被转化为硬脂酰辅酶A,再经去饱和作用形成油酰辅酶A。此外,酵母中的油酰辅酶A既能在线粒体或内质网中水解生成游离脂肪酸,也可以从游离脂肪酸重新酯化为油酰辅酶A。而在人体内,硬脂酰-ACP先水解生成游离脂肪酸,再合成硬脂酰辅酶A,最终通过去饱和作用形成油酰辅酶A。在神经酸生物合成的第2阶段,不同生物均以油酰辅酶A为底物,在脂肪酸延长酶(FAE)模块中的多种酶的循环催化下逐步延长碳链,但植物、酵母和人的FAE模块所涉及的酶类型存在一定差异,在图3b中以不同颜色标注。以植物体系为例,丙二酰辅酶A首先与酰基辅酶A经KCS催化缩合生成3-酮酰基辅酶A,并被3-酮酰基辅酶A还原酶(KCR)还原为3-羟基酰基辅酶A,再经3-羟基酰基辅酶A脱水酶(HCD)催化,脱去一分子水形成2,3-反式烯酰辅酶A;最后在反式-2,3-烯酰辅酶A还原酶(ECR)的作用下生成长链酰基辅酶A。每完成一次“缩合-还原-脱水-再还原”的循环,脂肪酸碳链即可延长两个碳原子,油酰辅酶A经3个循环后可生成神经酰辅酶A(C24:1),并且经过肯尼迪途径,神经酸以甘油三酯的形式储存在脂质体中。
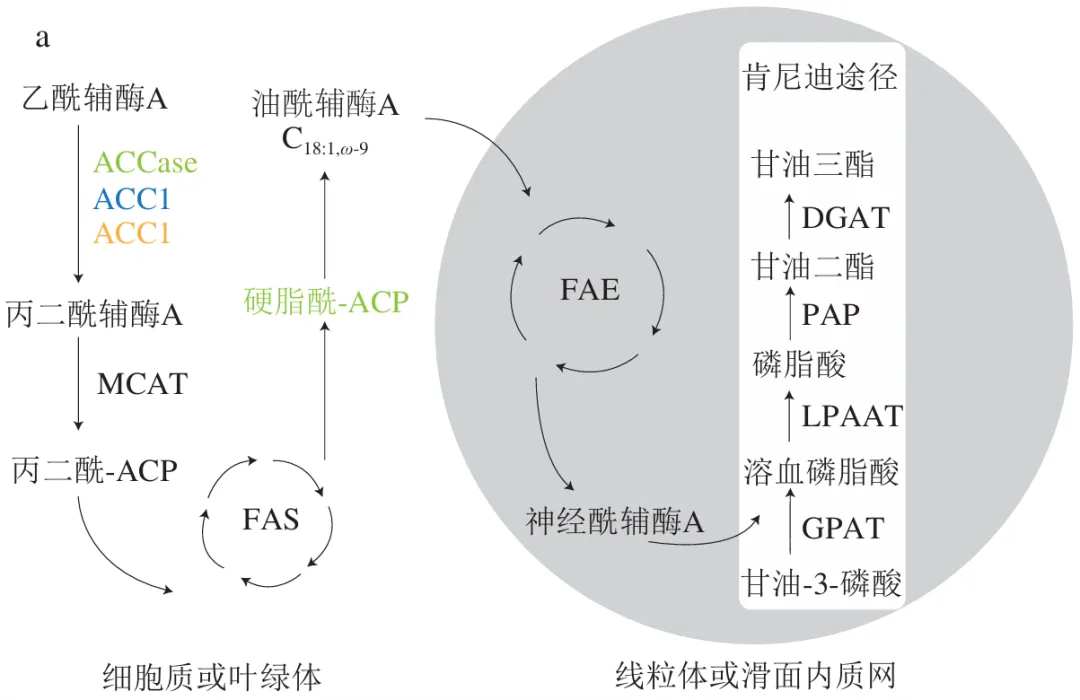 | 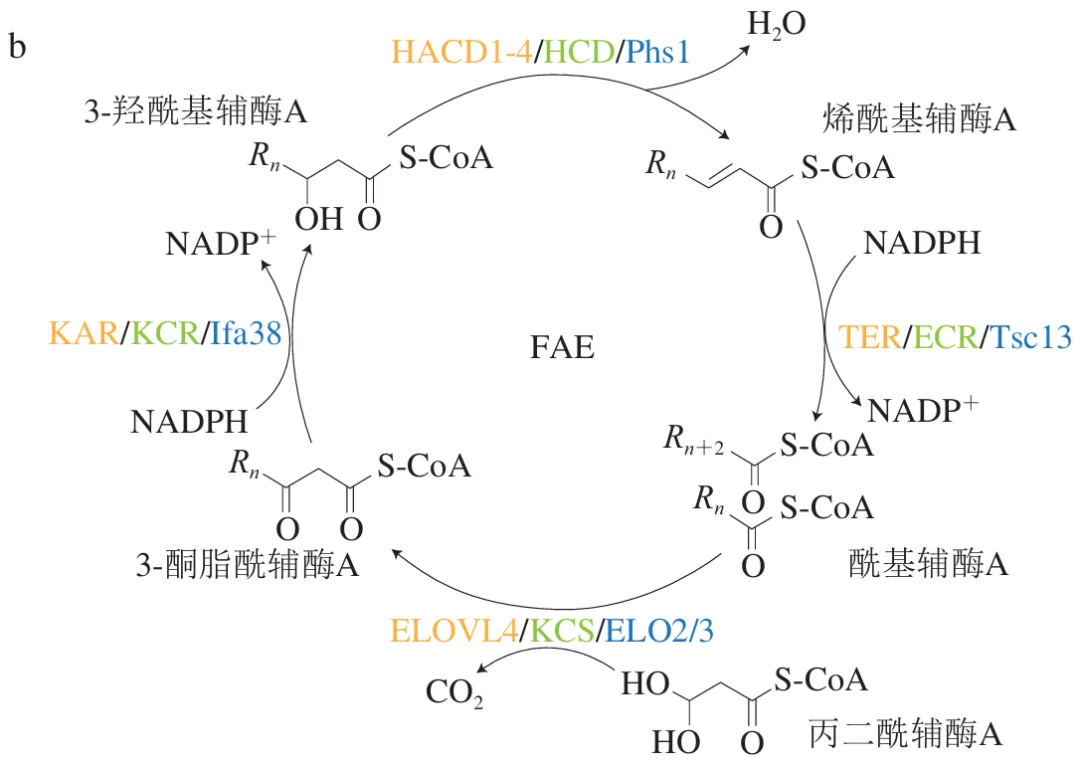 |

此外,研究表明不同来源的KCS具有显著的底物特异性,决定了脂肪酸碳链延伸长度。例如,Joubes等在拟南芥中鉴定出21个KCS基因,发现不同KCS基因在茎、叶、花及种子中的表达程度差异巨大,并可能影响超长链脂肪酸延伸长度。而Li Linkun等通过分析文冠果种子发育过程中神经酸含量的动态变化,推测XsKCS4、XsKCS7和XsKCS8这3种基因在神经酸生物合成中发挥重要作用。异源表达实验显示,XsKCS7在酵母与拟南芥中能够催化神经酸和芥酸(C22:1)的生物合成,但未表现出对C18:1的延长能力;而XsKCS4和XsKCS8则均未表现出C18:1、C20:1及C22:1的延长能力,证实了不同KCS在酰基辅酶A碳链延长过程中具有明确的底物选择性。近年来,通过代谢组学、转录组学以及核糖体足迹分析等手段,研究人员已系统解析了元宝枫、文冠果和蒜头果等植物的脂肪酸代谢特征及神经酸合成调控机制。这些研究不仅为阐明产油生物脂肪酸代谢网络奠定了理论基础,也为通过遗传与代谢工程优化底盘生物的神经酸合成能力提供了重要依据。
2.2 基因工程植物
如前所述,不同来源的KCS在酰基辅酶A碳链延长中表现出底物选择性差异。因此,在产油植物中异源表达特定的KCS基因,成为植物源神经酸生产的重要策略之一。如表3所示,将碎米荠的CgKCS基因导入高芥酸龙骨芥后,神经酸产量提高约15倍,芥酸占比也从未改造龙骨芥的38.0%~40.0%降低至5.0%~6.0%,表明CgKCS基因可能具有特异性延长芥酸碳链合成神经酸的能力。此外,银扇草富含芥酸和神经酸,其KCS基因(LaKCS)在龙骨芥中异源表达后,转基因植株籽油中神经酸占比可达30.0%。进一步分析表明,LaKCS的转录水平与芥酰辅酶A(C22:1)延长活性呈显著相关性,这证明该基因具有促进神经酸合成的能力。类似的,将蒜头果的MoKCS11基因在拟南芥中进行异源表达,可使拟南芥籽油中的神经酸占比从0.2%提高至5.0%,这表明蒜头果的KCS基因也具有强大的促进神经酸合成能力。然而,基因工程改造植物的周期较长,且在神经酸生产应用中仍面临土地占用量大、生长周期长、种子收集困难及芥酸含量高等限制,是植物法合成神经酸的主要瓶颈难题。

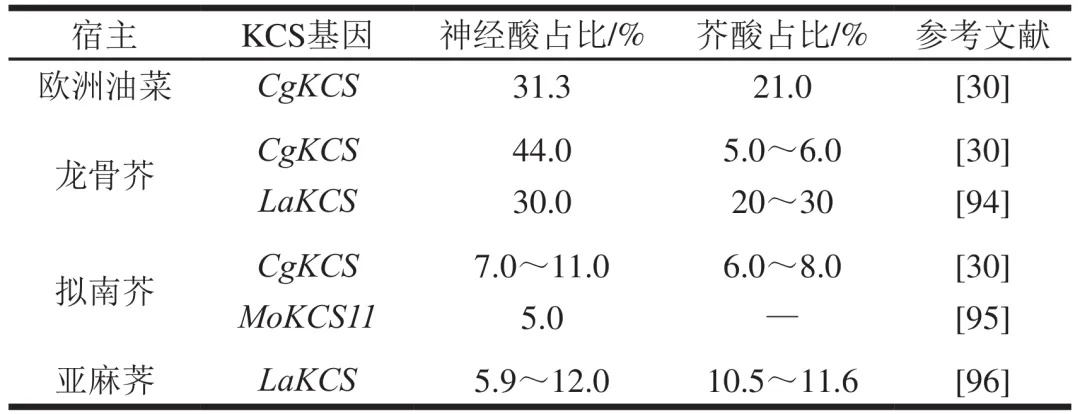
2.3 代谢工程改造酵母
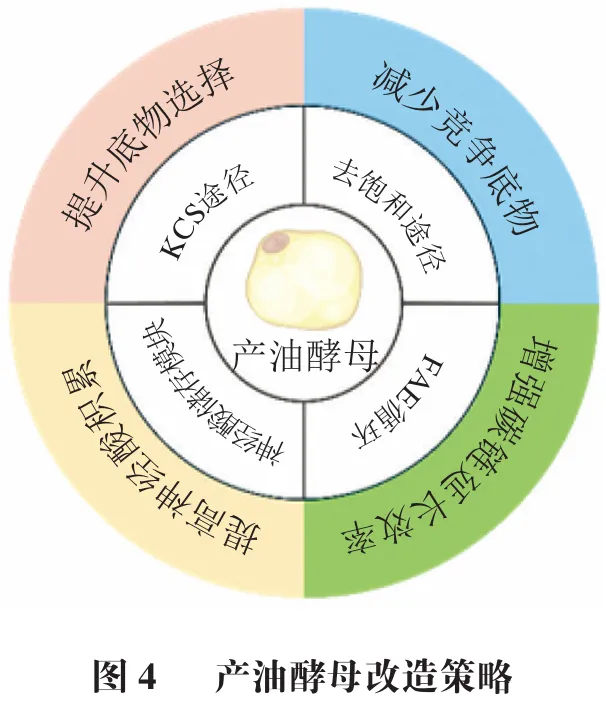
与土拉弗朗西斯菌等致病菌相比,酵母生物安全性更高、产油能力更强,已广泛应用于α-亚麻酸、二十二碳六烯酸(DHA)、二十碳五烯酸(EPA)等功能性脂肪酸的合成。早期改造微生物生产神经酸的策略相对简单,主要将高产神经酸植株的关键脂肪酸延长酶基因在产油酵母中异源表达提高神经酸生产效率。例如,Fillet等在高产油圆红冬孢酵母中共表达脂肪酸延长酶基因CgKCS(碎米荠)和CraKCS(海甘蓝),获得了总油脂滴度50.0 g/L、神经酸相对含量为16.0%的工程菌株。如表4所示,随着神经酸代谢途径的逐步阐明和发酵工艺优化,基于酿酒酵母、圆红冬孢酵母及解脂耶氏酵母等底盘菌株的神经酸生物合成改造策略正逐渐扩展。Liu Junfeng等利用β-雌二醇诱导表达系统增强脂肪酸延长酶基因(ELOVL1)的表达,并敲除了负责将C18:0延长至C24:0的ELO2基因,改造后的酿酒酵母中神经酸占比达到总脂肪酸的0.7%。Zhang Yue等通过在酿酒酵母共同表达MoKCS与ChFAE1并敲除ELO2,获得了神经酸占其总脂肪酸组成3.3%的工程菌株,并在优化培养条件后得到57 mg/L的神经酸滴度。这两项研究均采用过表达脂肪酸延伸酶基因,并结合对竞争途径的抑制策略对酿酒酵母进行改造,实现了神经酸在酵母中的合成积累,并且均验证了ELO2的缺失可有效削弱神经酸生物合成的竞争途径。

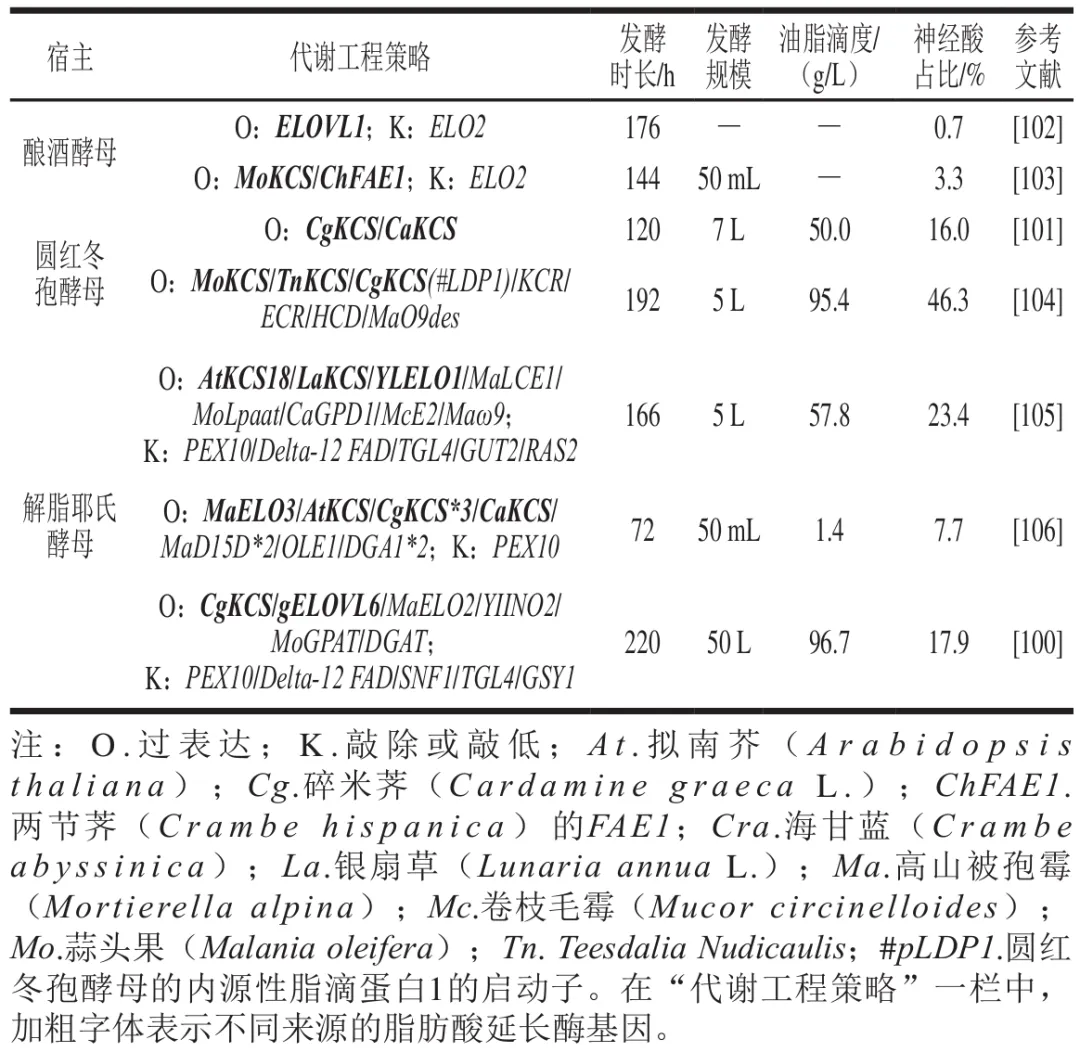
近年来,针对圆红冬孢酵母、解脂耶氏酵母等产油酵母神经酸生物合成策略的进一步完善和优化。Liu Feixiang等评估了3种植物来源KCS的催化偏好,并过表达圆红冬孢酵母内源KCR、ECR和HCD基因以增强FAE模块延长效率,同时利用启动子#LDP1强化CgKCS表达解决神经酸积累受限问题。在此基础上,通过表达去饱和酶基因MaO9des促进C24:0在n-9位形成双键,最终获得了一株油脂滴度95.4 g/L、神经酸在脂质组成占比46.3%的菌株。Wang Kaifeng等通过引入KCS基因和去饱和途径,结合β-氧化途径缺失,实现以“阻断-拉动-约束”的策略提高解脂耶氏酵母中硬脂酸前体的供应,并引入来自蒜头果的MoLpaat替换内源LPAAT,获得的菌株在5 L发酵罐中油脂生产达57.8 g/L,其中神经酸占比23.4%。Su Hang等在解脂耶氏酵母中反复表达编码CgKCS、gELOVL6和去饱和酶MaOLE2基因,并利用柔性甘氨酸连接体(GGGGGGGG)将MaOLE2与CgKCS偶联以提高油酰辅酶A利用率,结合内质网表达MoGPAT与DGAT、敲除AMP激活的蛋白激酶基因SNF1等手段,最终得到的解脂耶氏酵母在50 L发酵罐中神经酸滴度达17.3 g/L。
总体而言,改造产油酵母生物合成神经酸主要通过KCS基因组合表达、去饱和、FAE模块强化、神经酸储存优化等多重手段实现(图4)。针对不同底盘菌株的多策略代谢工程改造已被证明可有效提高神经酸的微生物合成水平。随着遗传操作工具的不断发展以及脂质代谢网络的进一步解析,预期将出现更多的通路构建与系统化改造办法,从而充分挖掘产油酵母或者其他底盘菌株的代谢潜力,再结合发酵工艺优化将大大促进神经酸的生物合成效率。
3
神经酸分离纯化
神经酸在动植物中含量普遍较低,而作为营养补充剂、药物和功能性食品配料时,产品纯度要求高,且芥酸占比需低于5%。因此,对含神经酸的粗品油进行分离纯化和富集精制,是提升其健康功效和应用场景的必要手段。此外,不同油脂提取工艺不仅会影响总油脂和神经酸得率,还会造成油脂理化性质、脂肪酸组成、营养成分及抗氧化能力差异。Zheng Yuling等对比冷榨法、正己烷萃取法、水酶法和超临界流体萃取法4种工艺提取含神经酸的文冠果油。发现冷榨法所得油脂的生育酚和甾醇含量最高,分别为530.4 mg/kg和2 105.1 mg/kg;而超临界流体萃取法所得油脂对1,1-二苯基-2-三硝基苯肼(DPPH)自由基的清除率最高。
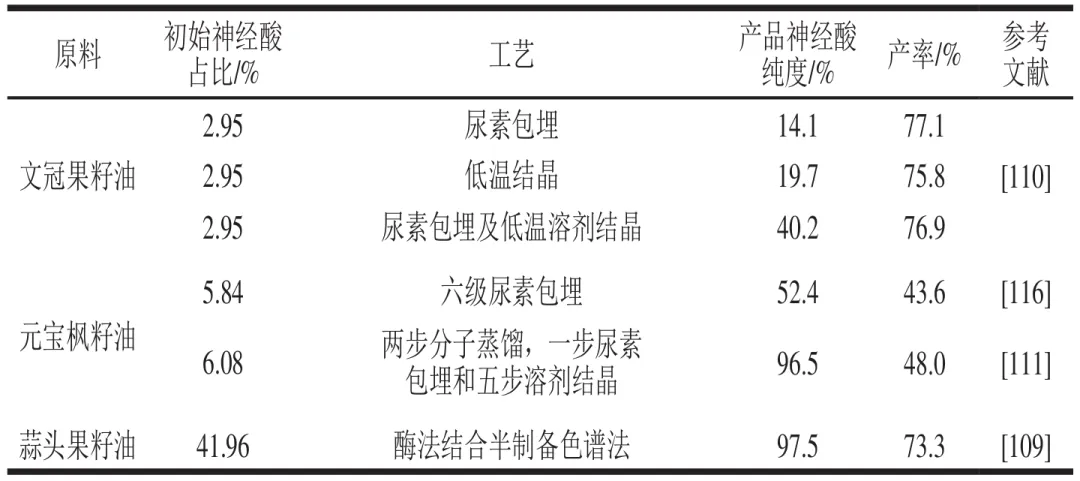
目前金属盐沉淀、尿素包合、低温结晶、分子蒸馏及减压精馏等方法已被应用于神经酸的分离纯化。但植物油脂中脂肪酸组成复杂,部分组分与神经酸结构相似,单一方法难以直接获得高纯度神经酸。如表5所示,Gao Hui等单独使用尿素络合法和低温溶剂结晶法对文冠果油进行纯化,所得产物神经酸纯度分别为14.1%和19.7%;但当两种方法联用时,神经酸纯度可大幅提升至40.2%。Gao Yingxi等采用分子蒸馏-尿素络合-溶剂结晶联合工艺纯化元宝枫籽油,经两级分子蒸馏去除棕榈酸(C16)和部分硬脂酸、油酸、亚油酸、亚麻酸(C18~C20),再通过尿素络合将C22~C24饱和脂肪酸占比分别降至0.2%和0.1%,最终结合溶剂结晶去除C18~C22不饱和脂肪酸,获得的神经酸纯度为96.5%,综合收率为48.0%。一般来说,不同纯化方法对脂肪酸的复杂组成具有一定特异性和成本差异。如分子蒸馏可优先快速去除短链脂肪酸,但处理量视设备规模而定,并且设备成本较高,真空要求提高了能耗成本。溶剂结晶设备需求简单且有较高处理量,可有效去除短链不饱和脂肪酸但可能导致饱和脂肪酸积累,对于油脂组成复杂神经酸含量较低的原料,溶剂结晶较适合于初步富集。而尿素包埋法设备投入相对较小,适用于大批量富集,可有效去除长链饱和脂肪酸,但若要达到高纯度仍需后工序。因此多方法联用可综合发挥不同方法的特异性,降低富集成本,但方法联用时需要针对不同方法处理后的组分变化考虑工艺顺序及参数调整。此外,目前神经酸的产业化发展仍处于起步阶段,这主要受限于天然植物中神经酸含量低、高纯度神经酸制备工艺复杂、成本高等因素。而酶法富集、乳化分离、柱层析及色谱等方法已在DHA、EPA等多不饱和脂肪酸的富集精制和产品创制中得到广泛应用,可为高纯度神经酸制取提供借鉴。

4
神经酸健康功效及产品开发
4.1 健康功效
神经酸在人体神经系统和脂质代谢中发挥重要作用,其在人体内的代谢水平一定程度上反映了人体的健康情况。如Kunst等指出,肠炎患者粪便中神经酸水平随炎症标志物粪便钙卫蛋白水平升高而增加,并且接受皮质类固醇或白介素-12/23抗体治疗的患者,其粪便神经酸水平高于其他治疗组,这显示粪便神经酸含量或可用于肠炎诊断及治疗进展评估。此外,Dhillon等发现在轻度认知障碍人群中,简易精神状态检查评分与血红细胞中神经酸水平呈正相关(r=0.54,P=0.01),即血红细胞脂肪酸组成中神经酸浓度变化可能用于痴呆风险评估。这些研究表明,内源性神经酸水平与脂质代谢紊乱及神经系统疾病密切相关,有望作为相关疾病诊断、进展评估及治疗监测的特异性生物标志物。
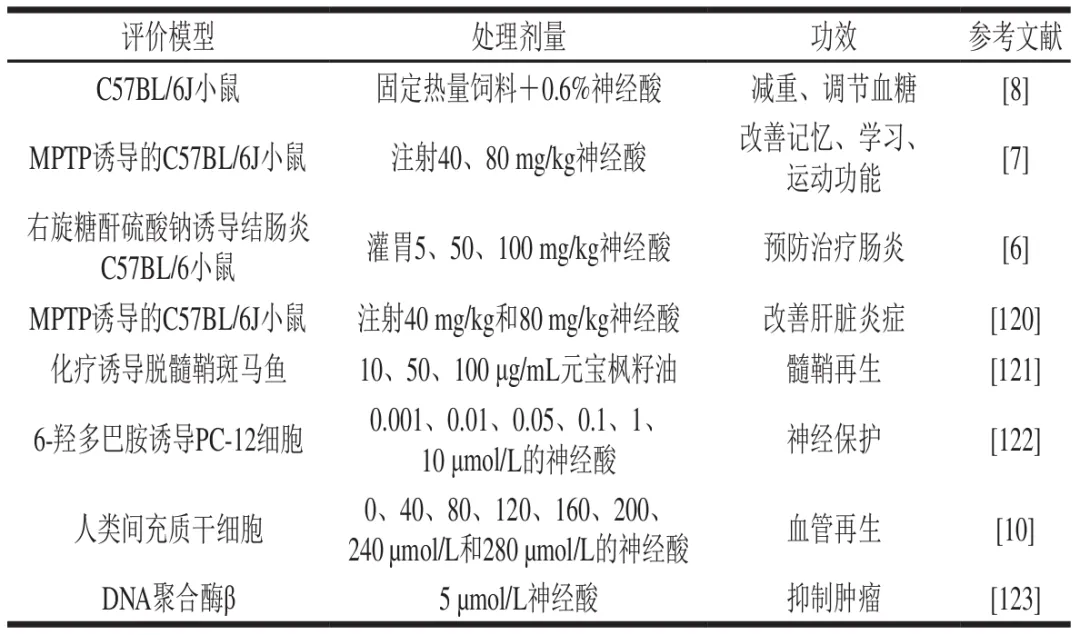
神经酸除上述生物标志物的应用潜力外,还具有保护与修复神经、促进脑发育、提升认知功能等多种生理功效(表6)。例如,通过在高脂喂养小鼠模型的饲料中添加0.6%神经酸,不仅可以调节脂质代谢水平降低小鼠体质量,还可以有效控制小鼠血糖。Wang Xueqi等则发现对1-甲基-4-苯基-1,2,3,6-四氢吡啶(MPTP)诱导的C57BL/6J小鼠连续10 d腹腔注射神经酸可增强突触可塑性、减弱神经炎症、提高小鼠爬杆和迷宫测试中运动及认知表现、改善记忆与学习功能。而该团队使用同样的模型和注射给药策略,发现神经酸还可通过抑制炎症因子的信号通路,显著改善小鼠肝脏炎症。类似地,对小鼠灌胃神经酸同样可以抑制炎症因子水平及炎症信号通路,并可调节多种代谢途径发挥抗炎作用。Cheng Dekun等发现100 μg/mL的元宝枫籽油可以逆转脱髓鞘斑马鱼尾部形态损伤、改善轴突退化和触觉反应延迟等功能,并能降低髓鞘基础蛋白表达,改善线粒体功能障碍促进髓鞘再生。Umemoto等则发现使用极低剂量的神经酸(0.001~0.1 μmol/L)预处理48 h,即可缓解6-羟多巴胺诱导PC-12细胞的氧化应激损伤、提高细胞存活率,对神经细胞发挥保护作用。而在人类间充质干细胞培养基中引入神经酸,则可以增加血管生成相关基因的表达,从而促进血管再生。此外,神经酸还可以通过减弱DNA聚合酶β与双链DNA结合能力,抑制肿瘤细胞生长。这些研究表明神经酸在肥胖、糖尿病、阿尔茨海默病和癌症等疾病的治疗中发挥积极作用。然而,大多数研究仍停留在细胞和特定动物模型,缺乏系统的临床数据支持,关于其疗效、使用剂量及长期安全性仍需要验证评估。其治疗潜力的发掘,还需要明确其在神经传递、炎症及代谢调控中的具体分子通路。

4.2 产品开发
得益于神经酸特有的健康功能及理化特性,为满足不同人群和应用场景,目前神经酸产品开发主要集中于食用油、微囊粉、乳剂、功能结构脂、饮料等营养强化食品(表7)。首先,随着国家对木本植物油的推广,以神经酸为主要营养素的元宝枫、文冠果及蒜头果等作物受到广泛关注,其中元宝枫籽油国家标准(GB/T 37748—2019《元宝枫籽油》)的实施,对推动神经酸在食品和医药产业中的应用具有重要意义。然而,神经酸作为一种长链单不饱和脂肪酸,在光照、高温、金属离子等因素作用下其不饱和双键极易发生氧化,导致共轭二烯烃、氢过氧化物、醛酮化合物等氧化产物生成。这些氧化产物不仅导致神经酸油脂产生哈喇味、营养价值下降,甚至严重威胁人体健康安全。


研究人员开发了乳剂、软胶囊、微囊粉等油脂包覆技术,不仅可以有效提升神经酸氧化稳定性,还可以增强其水溶性和生物利用度。例如,陈文静等使用神经酸、油酸、辛基苯酚聚氧乙烯醚-10及丙二醇为原料,在高压均质条件下获得平均粒径约158 nm、外观均一且稳定性良好的乳剂,该制剂在室温长期存放6个月仍能保持稳定。Jin Yu等则使用1.5%大豆分离蛋白作为乳化剂,和0.5 mg/mL的神经酸在120 MPa高压均质制备得到平均粒径处于纳米尺度的稳定乳剂,在热处理、4 ℃储存30 d等实验条件下均表现出良好稳定性。Li Yanmei等采用亚临界流体提取含有神经酸的羊栖菜藻油,并通过连续凝聚包埋创制水溶性抗氧化微胶囊,该产品具有抑制氧化应激与预防神经炎症的功能,并在斑马鱼及小鼠模型中表现出显著的记忆增强作用。而Xu Xiaojie等采用反溶剂法将带负电的神经酸与带正电的壳聚糖结合,并通过带负电的丝素蛋白对壳聚糖-神经酸复合物进行包封,制备了2.5 h内神经酸释放达89.5%的水溶性复合微囊粉,该产品能够通过抑制氧化应激和降低线粒体膜电位保护神经细胞。此外,神经酸是母乳中的天然成分,其含量会随哺乳期的延长而逐渐下降,因此在婴幼儿配方奶粉中添加神经酸时,需要考虑精确调整脂肪酸组成,以模拟母乳特征满足婴儿个体化营养需求。这些研究为神经酸营养强化剂和配方产品的开发奠定了基础,拓展了其在孕产妇、学龄儿童、老年受众等特定人群的应用前景。
除了高纯度的神经酸原料外,开发结构脂质如含神经酸中长链甘油三酯及结构磷脂等也是未来发展的重要方向。例如,Lai Yundong等使用自制的固定化脂肪酶CSL@HHSS,催化蒜头果油与中链甘油三酯发生酯交换反应,成功制备了含神经酸中长链甘油三酯(占总甘油三酯的62.3%)。该产品结合了中链甘油三酯和长链甘油三酯的特性,协同了神经酸的健康功效及中链脂肪酸迅速吸收和代谢的特点,促进神经酸生物利用效率。Ang Xun等采用固定化磷脂酶A1催化神经酸甲酯与磷脂酰胆碱反应,在最优条件下(55 ℃、水添加量1%、反应9 h),制备的磷脂酰胆碱产品神经酸占其总脂肪酸的48.2%。作为细胞膜和髓鞘的重要组成部分,磷脂酰胆碱可高效地运送神经酸至神经系统,实现靶向递送。这些新型功能酯可能在营养保健与医学应用中增强神经酸的生物学功效,但具体生物活性及脂质功能变化仍需通过动物模型和体外实验进行验证。此外,神经酸及其衍生物还均未获得GRAS认证,并未被各国际组织及国家列为食品添加剂。未来对于神经酸产品开发还需要进一步开展临床试验、安全性评估、口感接受度评价及推荐剂量确认等研究,并在此基础上推动法规及标准完善,争取列入新的食品、特殊医学用途配方食品原料。
5
结 语
神经酸具有神经保护、血糖调节、抗炎作用及认知功能改善等功能,在预防和治疗神经系统疾病、癌症、肥胖、糖尿病等疾病方面展现出治疗潜力。目前,商业来源以元宝枫、文冠果等植物种植为主,但存在种植周期长、亩产有限等可持续发展难题。以解脂耶氏酵母、圆红冬孢酵母等微生物合成神经酸的技术已取得突破进展,结合KCS基因组合表达、去饱和策略、FAE模块强化等系统性代谢改造和发酵工艺优化,可提升生产效率和产量,减少耕地占用和农药使用。此外,神经酸的富集精制仍是其产品开发应用的关键步骤,可借鉴DHA、EPA等产业化的功能性脂肪酸的纯化工艺,结合含神经酸油脂的脂肪酸组成特性,联用多种绿色纯化工艺如酶法前处理及色谱技术等可制备高纯度的神经酸产品。在此基础上,使用油脂包覆技术可提升神经酸氧化稳定性、增强水溶性和生物利用度,开发磷脂型、甘油酯性等神经酸结构脂,可提高吸收利用率、协同增强神经酸健康功效,这些技术为神经酸产品创制提供了新思路。随着神经酸生产成本降低、功效剂量确认、安全性稳定性评估及相关法规标准的完善,未来神经酸将会在食品营养补充剂、保健食品和医药等领域展现广阔的应用前景。
作者简介



实习编辑:杨瑞蕾;责任编辑:张睿梅。点击下方阅读原文即可查看全文。图片来源于文章原文及摄图网
近期研究热点
《食品科学》:华南农业大学肖杰教授等:大数据与机器学习赋能功能食品稳态增效递送体系的智能化构建前沿进展
《食品科学》:陕西科技大学姚晓琳教授等:口服胶原蛋白水解物的抗皮肤光老化机制及促吸收策略研究进展
《食品科学》:昆明理工大学张付杰副教授等:基于近红外光谱技术结合机器学习的咖啡粉掺假检测分析
《食品科学》:北京三元食品股份有限公司陈历俊教授级高级工程师等:基于UPLC-QTOF-MS技术的母乳非靶向代谢组学分析方法的建立与优化
《食品科学》:四川轻化工大学黄治国教授等:不同质量浓香型白酒窖泥细菌群落结构差异性分析
《食品科学》:哈尔滨商业大学张娜教授等:不同盐离子与淀粉间的互作及其对淀粉理化性质的影响研究进展
《食品科学》:河南工业大学郑学玲教授等:小麦系统粉淀粉结构差异对其理化特性的影响




长按或微信扫码进行注册
为对标农业农村部2035年科技规划及“十四五”“十五五”发展方向,推动农产品加工与储运的工程化、智能化、绿色化升级,由湖南省农业科学院、湖南农业大学、北京食品科学研究院、国际食品科技联盟(IUFoST)、中国农业大学、岳麓山工业创新中心主办,湖南大学、中南林业科技大学、长沙理工大学、湖南中医药大学、湘潭大学、岳麓山实验室协办,中国食品杂志社、洞庭实验室、湖南省食品科学技术学会、湖南省农产品加工与质量安全研究所、湖南农业大学食品科学技术学院、Springer Nature-《Agricultural Products Processing and Storage》杂志承办的“第二届农产品加工与食品制造国际学术研讨会—创新引领绿色智造,AI赋能科技进步”,将于2026年9月19-20日(9月18日会议报到)在中国 湖南 长沙召开。

长按或微信扫码进行注册
会议招商招展
联系人:杨红;电话:010-83152138;手机:13522179918(微信同号)

随机文章
-
 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
- 琼中科协召开农业领域科技工作者座谈会 凝聚科技力量服务“三农”发展
- 农业农村部公告!仙游2个产品入选
- 我家遭遇洪水农作物全毁了,临时救助和农业保险能同时申请吗?
- 事业编制 | 中国热带农业科学院环境与植物保护研究所2026年度第二批公开招聘工作人员公告
- 会宁县科技服务“组合拳”赋能旱作农业
- 【招聘公告】新疆农业职业技术大学2026年第一批面向社会引进博士研究生公告
- 跨域携手赋能数字现代农业|云桂通科技联袂河北凌寒科技达成战略合作,共建佳慧甄选平台绿色认耕代养数字平台
- 深化产学研协同 赋能生态农业落地|南京首建投组织农业专项培训 特邀锐昌生态储海岩分享原创土壤改良技术
- 农业农村领域有限空间作业安全指南请收好!
- 区畜牧渔业中心第22周工作动态
