2026年,中国农业大学植物保护学院农林生物安全全国重点实验室周欣、罗诗琦研究团队在《Nature Communications》期刊上发表了题为“Gut microbiota-derived butyrate primes systemic immunity in honey bees by mediating lipid metabolic reprogramming”的文章。该研究以蜜蜂为模式生物,整合宏基因组学、RNA-seq转录组测序、LC-HRMS非靶向代谢组学、CUT&Tag H3K27ac表观遗传分析以及Western blot、PGE2 ELISA等生化手段,系统解析了肠道核心菌群通过代谢物丁酸盐远程调控宿主系统性免疫的分子机制,首次揭示了"丁酸盐→GPR41/HDAC双通路→脂肪体脂质代谢重编程→前列腺素E2(PGE2)生物合成→体液与细胞免疫激活"的完整信号轴,为理解肠道-宿主免疫-代谢互作提供了全新视角,并为利用益生菌或代谢物干预改善传粉昆虫健康奠定了理论基础。
昆虫虽不具备脊椎动物那样的适应性免疫,却能通过"免疫启动"(immune priming)机制增强对后续病原体感染的抵抗力——这一现象功能上类似于脊椎动物的"免疫记忆"。蜜蜂作为全球最重要的传粉昆虫之一,其肠道微生物群组成高度保守且稳定,以5个核心细菌属为优势类群:Gilliamella、Snodgrassella、Bifidobacterium、Bombilactobacillus 和 Lactobacillus。先前研究已证实,肠道菌群定植能增强蜜蜂的抗病能力,但活菌的保护效应显著优于热灭活细菌,提示细菌代谢产物在此过程中发挥关键作用。然而,肠道菌群如何远程调控脂肪体(昆虫重要的代谢与免疫器官)和血淋巴中的系统性免疫,其分子机制长期不清楚。
1.丁酸盐是蜜蜂肠道菌群启动宿主系统性免疫的关键代谢分子。
2.丁酸盐通过激活GPR41信号通路和抑制HDAC活性双重机制调控脂肪体脂质代谢基因表达。
3.PGE2是连接脂质代谢重编程与免疫激活的关键下游信号分子。
4.丁酸盐通过上调H3K27ac表观遗传修饰重塑脂肪体代谢基因染色质状态。
1. 活菌通过代谢产物远程激活系统性体液与细胞免疫
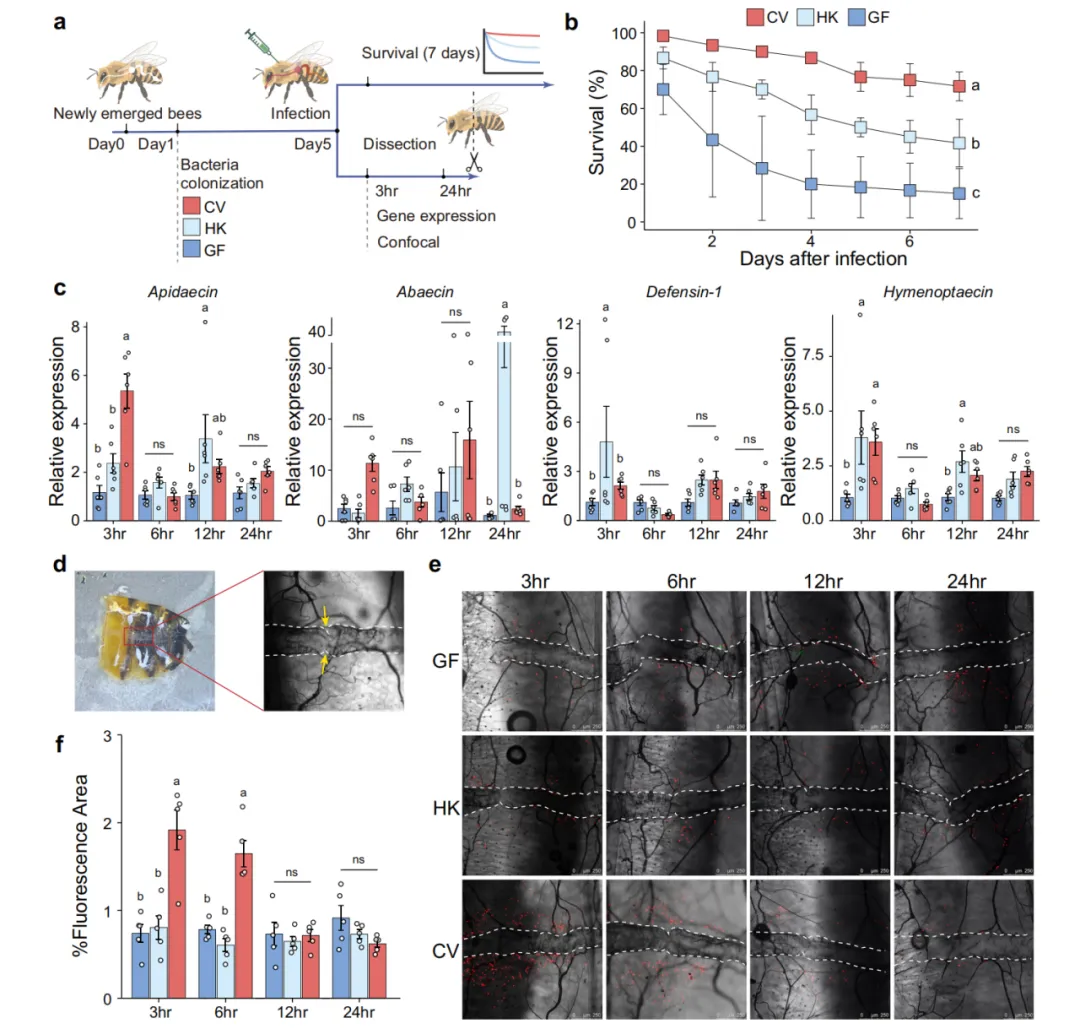
研究团队首先构建了无菌(GF)、热灭活菌群(HK)和正常菌群定植(CV)三组蜜蜂,以条件致病菌 Hafnia alvei 腹腔注射作为感染模型。生存曲线分析表明,CV组蜜蜂存活率显著高于GF和HK组,提示活菌代谢活性而非菌体成分是保护效应的关键介导因子。qPCR检测脂肪体中抗菌肽(AMP)基因表达,结果显示CV组蜜蜂的 Apidaecin、Abaecin、Hymenoptaecin 和 Defensin-1 表达量均显著高于GF组,证实肠道菌群可远程激活脂肪体这一关键代谢-免疫器官的体液免疫输出。细胞免疫层面,围心室血细胞聚集程度在CV组中显著增强(约2.6倍),而GF和HK组几乎无聚集反应,提示肠道菌群对昆虫细胞免疫的远程调控作用(图1)。
2. 丁酸盐——肠道菌群免疫启动效应背后的关键代谢物
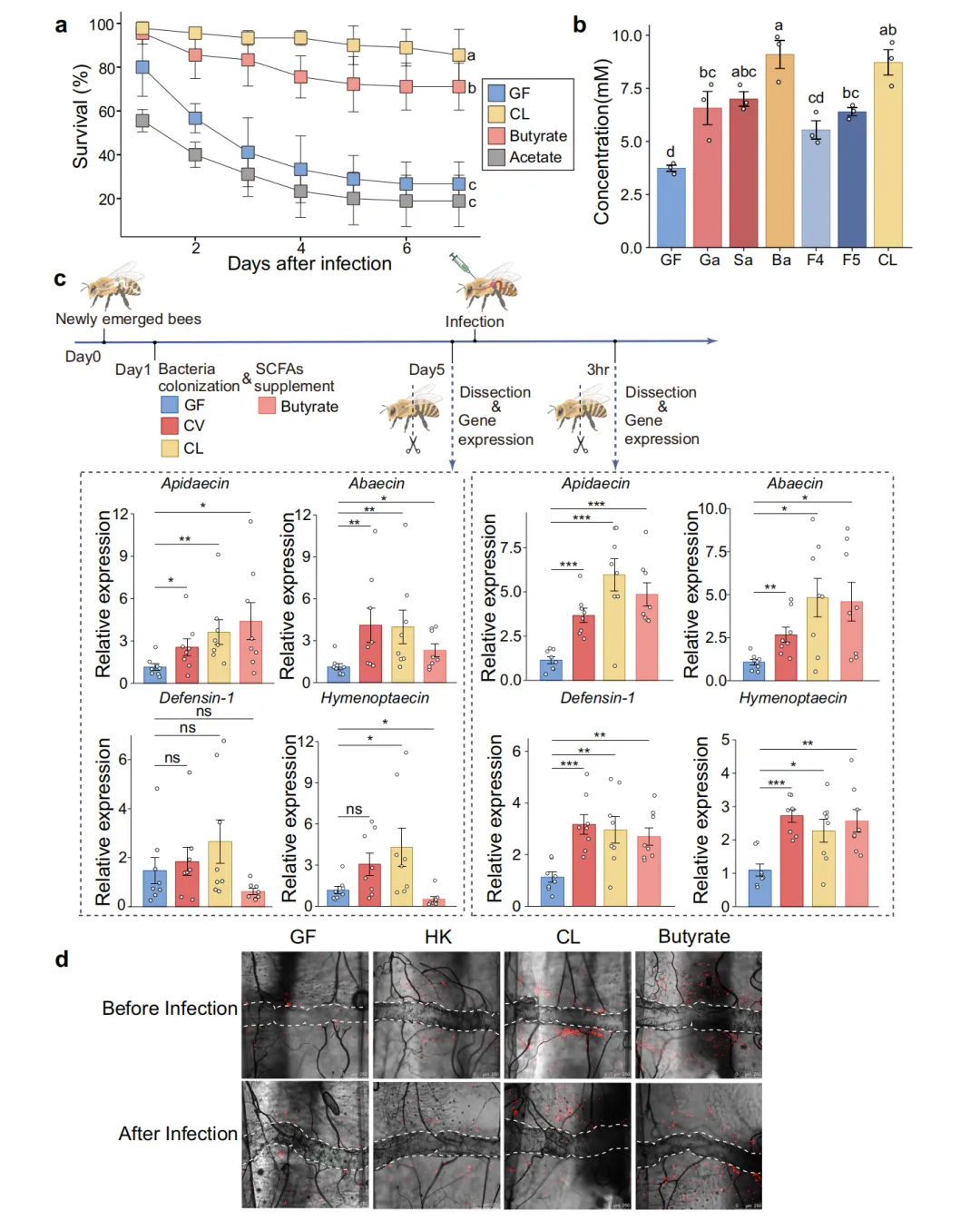
为鉴定活菌中发挥保护作用的具体代谢物,研究团队对GF蜜蜂分别补充了5种核心菌的混合培养物(CL)、醋酸盐或丁酸盐。生存率分析显示,丁酸盐补充组蜜蜂存活率与CL组相当,而醋酸盐补充组无保护效应。单菌株定植实验的腹部丁酸盐定量结果进一步揭示,Bifidobacterium 定植组的丁酸盐产量最高,与生存率数据高度吻合,提示特定菌株通过产丁酸盐发挥免疫激活功能。qPCR检测脂肪体AMP表达证实,丁酸盐补充能恢复GF蜜蜂在 Hafnia 感染前后AMP的表达水平。围心室血细胞聚集荧光成像进一步表明,丁酸盐处理能恢复GF蜜蜂的血细胞聚集能力,而HK和CL组间无显著差异(图2)。
图 2 蜜蜂肠道细菌产生的丁酸盐启动宿主系统性免疫
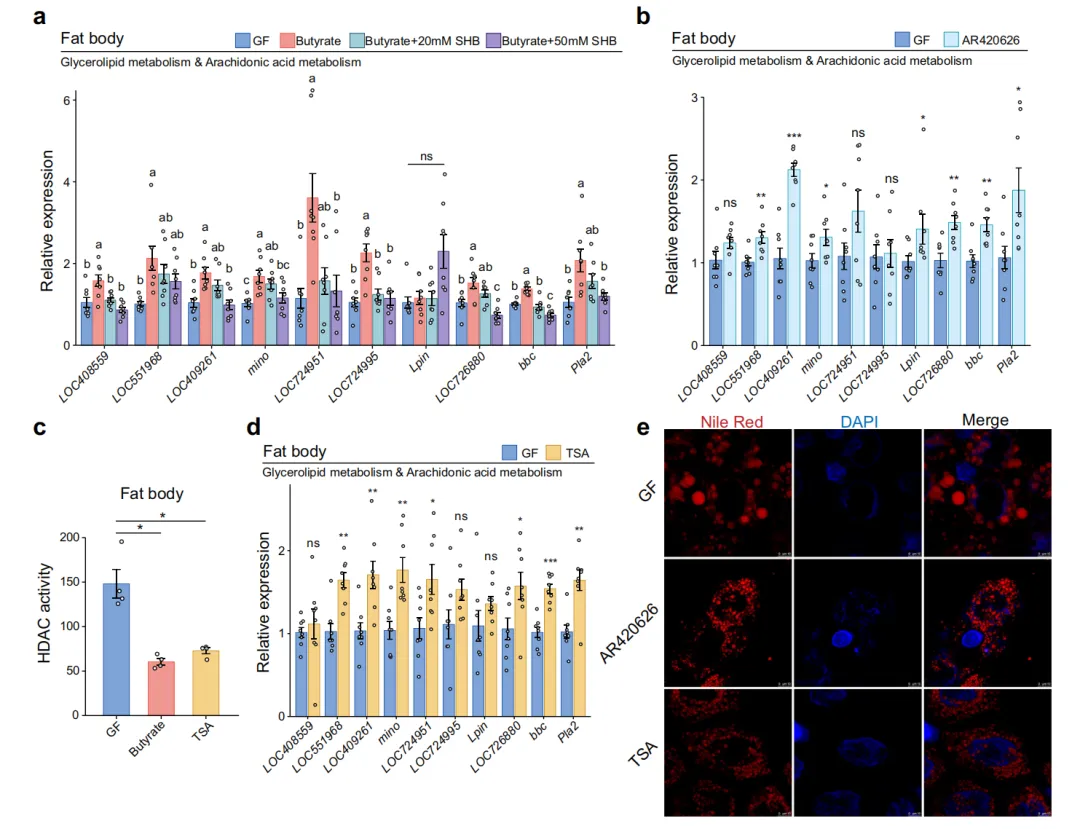
3. 丁酸盐重编程脂肪体脂质代谢:以花生四烯酸-PGE2通路为核心
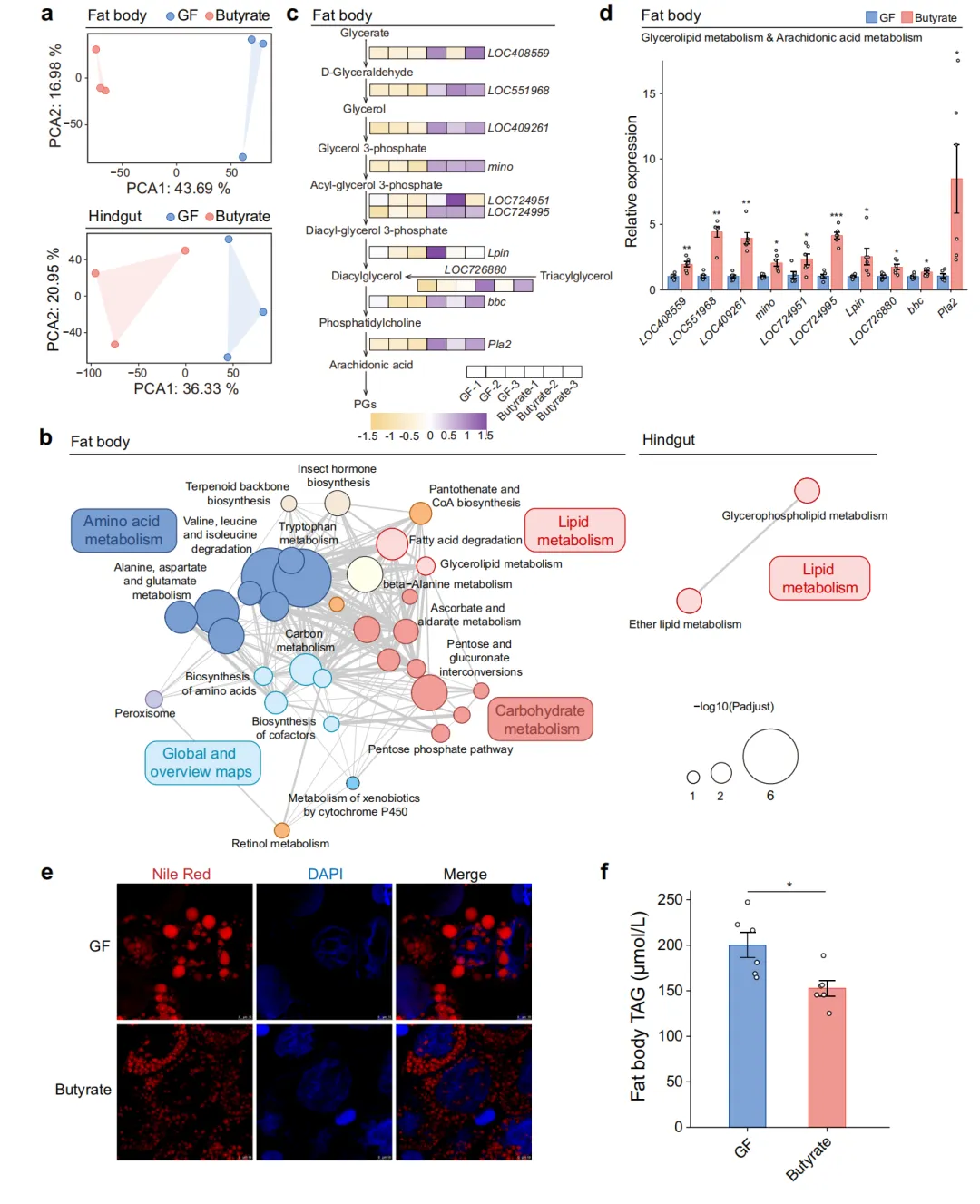
为解析丁酸盐调控免疫的分子机制,研究团队对GF和丁酸盐补充组蜜蜂的脂肪体与后肠进行了RNA-seq分析。主成分分析(PCA)显示两组样本在转录组层面存在显著差异,表明丁酸盐引发了深度的基因表达重编程。通路富集分析以GF为对照,发现丁酸盐处理后脂肪体和后肠中上调的基因富集于甘油脂质代谢(Lpin、Pla2、mino 等)和花生四烯酸代谢通路。热图展示了参与脂质代谢的关键差异表达基因,包括视黄醛脱氢酶1、甘油激酶、1-脂酰甘油-3-磷酸酰基转移酶(LOC724951、LOC724995)、磷脂酸磷酸酶(Lpin)和磷脂酶A2(Pla2)等。qPCR在独立样本中对代表性基因进行了验证。Nile Red中性脂质染色显示丁酸盐处理5天后脂肪体中性脂质(主要是三酰甘油,TAG)显著降低,与脂肪酸氧化增强的代谢表型一致(图3)。
图 3 丁酸盐增强蜜蜂脂质分解,生成花生四烯酸代谢底物
4. PGE2——连接脂质代谢与免疫激活的关键信使
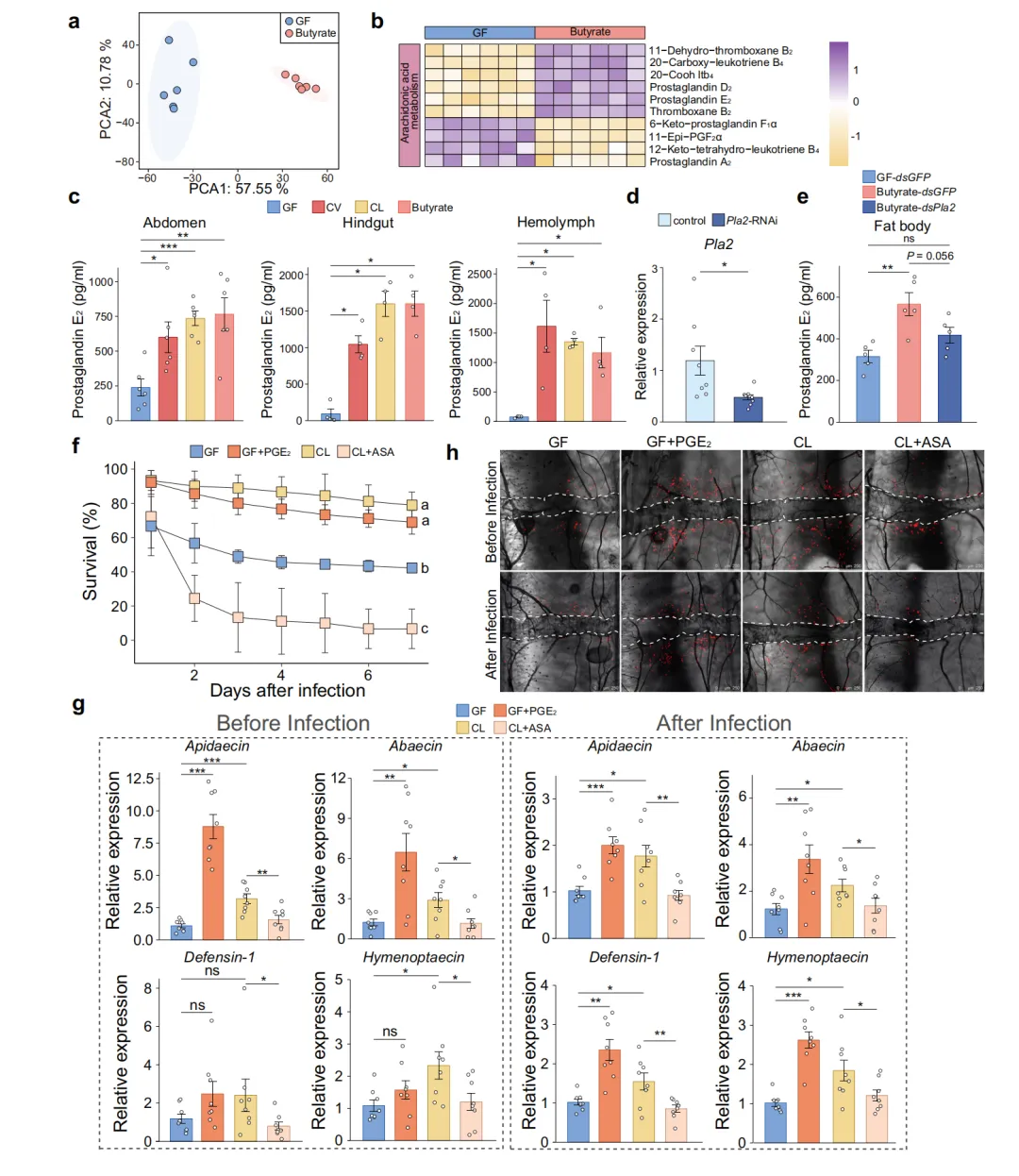
花生四烯酸代谢的核心产物前列腺素E2(PGE2)是哺乳动物炎症与免疫反应的重要调节因子,研究团队推测PGE2可能同样介导了蜜蜂的免疫启动效应。腹部代谢组PCA显示GF和丁酸盐组间存在显著代谢差异。花生四烯酸通路代谢物热图进一步揭示丁酸盐处理后PGE2及其前体水平升高。ELISA检测多个组织中的PGE2浓度,发现丁酸盐补充和菌群定植组蜜蜂的腹部、后肠和血淋巴中PGE2均显著高于GF组。通过RNAi敲降脂肪体 Pla2(PGE2合成的限速酶),PGE2水平显著降低;而丁酸盐对PGE2的上调效应在 Pla2 敲降后消失,证实Pla2是丁酸盐→PGE2通路中的必要节点。功能实验显示,外源PGE2补充恢复了GF蜜蜂的存活率;反之,ASA(前列腺素合成抑制剂)则削弱了CL组蜜蜂的保护效应。PGE2处理还恢复了GF蜜蜂的AMP表达和血细胞聚集能力,在分子和细胞两个层面验证了PGE2的下游免疫效应(图4)。
5. GPR41信号通路与HDAC抑制——丁酸盐的双重分子靶点
丁酸盐在哺乳动物中已被广泛报道可同时作为GPR41/43配体和HDAC抑制剂,研究团队接下来验证这两条机制在蜜蜂中的贡献。β-羟基丁酸盐(βHB,GPR41激动剂)处理能上调脂肪体中甘油脂质代谢和花生四烯酸代谢基因的表达,模拟了丁酸盐的部分效应。GPR41拮抗剂AR420626处理则完全阻断了丁酸盐对代谢基因的上调,证实GPR41通路在丁酸盐信号传导中不可或缺。HDAC酶活性检测显示,丁酸盐和TSA(HDAC抑制剂)处理均显著降低了脂肪体HDAC活性。TSA处理同样上调了脂质代谢基因表达,且与丁酸盐处理效果相当,提示HDAC抑制也参与了丁酸盐的转录调控。Nile Red染色证实AR420626和TSA处理均逆转或模拟了丁酸盐对脂肪体中性脂质水平的调节效应。这些数据共同表明,丁酸盐通过GPR41信号激活和HDAC抑制双重机制协调蜜蜂的脂质代谢-免疫基因表达(图5)。
图 5 丁酸盐通过激活GPCR通路并调控组蛋白乙酰化,平衡脂质代谢与免疫激活
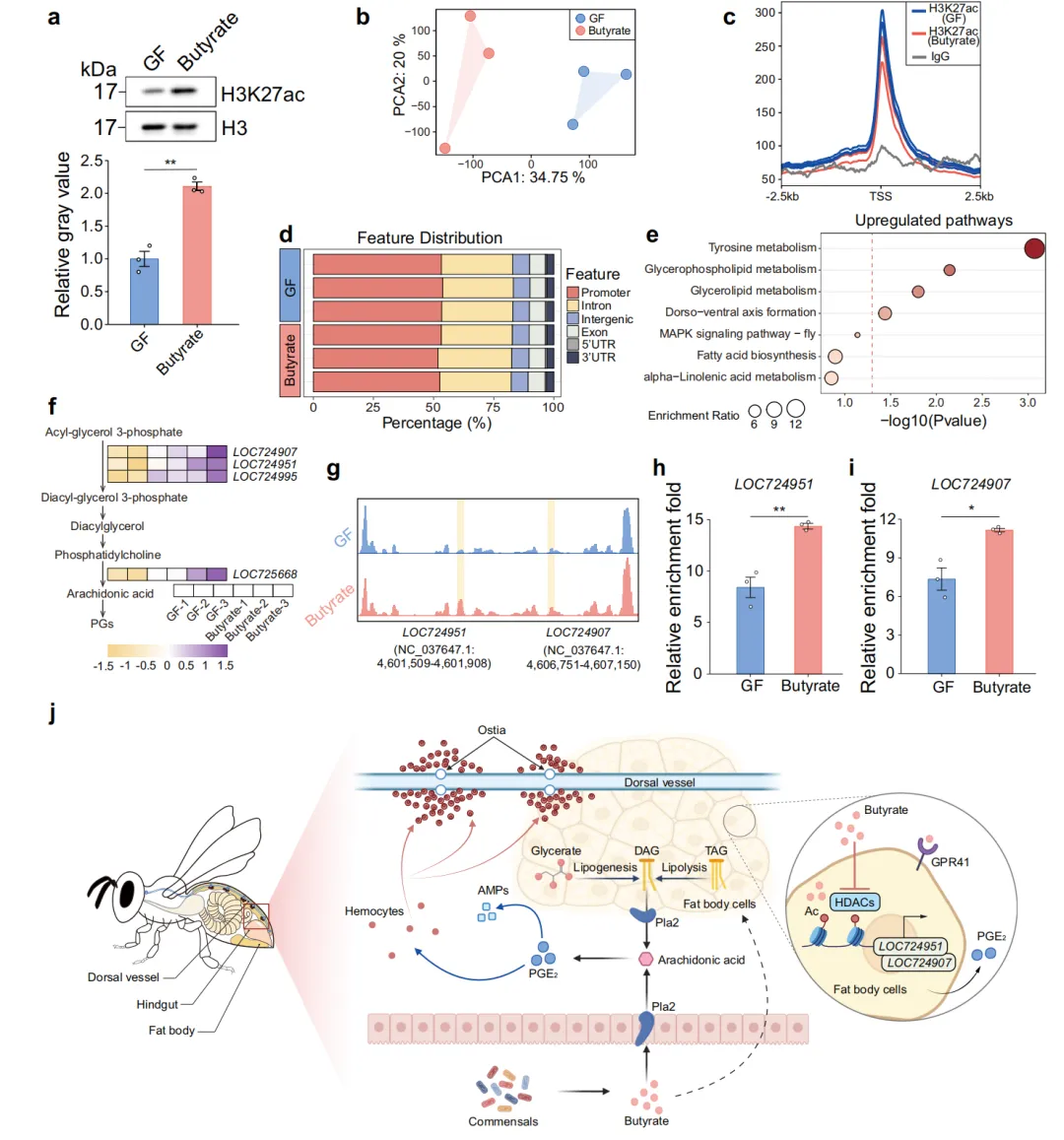
6. H3K27ac表观遗传修饰——丁酸盐长效调控脂质代谢的分子记忆机制
HDAC抑制与组蛋白乙酰化修饰密切相关,研究团队进一步探究丁酸盐是否通过表观遗传重塑来维持对代谢基因的长效调控。Western blot检测显示,丁酸盐处理5天后脂肪体H3K27ac全局水平显著升高(H3为上样内参),表明染色质处于更开放的状态。CUT&Tag H3K27ac测序的PCA分析进一步证实GF和丁酸盐组在表观遗传层面的显著差异。两组H3K27ac峰的overlap分析展示了各自特有的修饰区域。H3K27ac峰在基因组功能元件上的分布分析揭示了启动子区修饰的富集变化。KEGG通路分析表明,H3K27ac差异峰关联基因富集于甘油磷脂代谢通路。热图展示了甘油磷脂代谢基因上的H3K27ac差异修饰模式。IGV基因组浏览器track图展示了 LOC724951 和 LOC724907 两个代表性基因上H3K27ac信号的显著增强。ChIP-qPCR在两个基因启动子区验证了H3K27ac富集的显著增加(P = 0.0044和P = 0.0122),从遗传修饰层面确证了丁酸盐→HDAC抑制→H3K27ac增加→代谢基因激活的表观遗传调控链条。研究最终提出了一个完整的机制模型:肠道 Bifidobacterium 等菌株产生丁酸盐,通过GPR41信号通路和HDAC抑制(导致H3K27ac增加)双重机制重编程脂肪体脂质代谢,上调花生四烯酸→PGE2生物合成通路,最终启动体液(AMP)和细胞(血细胞聚集)免疫应答(图6)。
图 6 丁酸盐通过H3K27ac修饰调控蜜蜂脂肪体脂质代谢
本研究以蜜蜂为模式,首次系统揭示了肠道核心菌群通过丁酸盐远程调控宿主系统性免疫的完整分子机制,建立了"肠道菌群→丁酸盐→GPR41/HDAC双通路→脂肪体H3K27ac表观遗传重塑→脂质代谢重编程→PGE2生物合成→体液与细胞免疫激活"的信号轴。这一机制模型的建立不仅填补了昆虫免疫启动领域"肠道-代谢-免疫"跨器官调控的知识空白,更为难培养昆虫或野生动物中肠道菌群如何系统性影响宿主抗病力提供了可借鉴的研究范式。值得注意的是,本研究虽然以西方蜜蜂为对象,但其核心信号通路与哺乳动物共享——丁酸盐同样在人体中由肠道菌群产生并通过GPR41/HDAC调控代谢与免疫,提示这种"菌群代谢物-脂质代谢-PGE2-免疫"的调控逻辑可能是一种跨物种的高度保守机制,为比较免疫学研究提供了独特视角。
中国农业大学植物保护学院罗诗琦副教授为论文通讯作者,博士研究生刘佳铭为第一作者,周欣教授为论文做出了重要贡献。已毕业的硕士研究生吴亚帅、博士研究生李振芳、已毕业的博士研究生唐浚博参与了该项工作。本研究得到了科技部重点研发项目,国家自然科学基金和中国农业大学2115人才培育发展支持计划的资助。
微信:Metware-plant
电话:400 100 8862
邮箱:support@metware.cn
网址:www.metware.cn
点击文末“阅读原文”即可下载原文文献
我知道你在看哟














 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
