Microbiome | 中国热带农业科学院:链霉菌诱导的3-磷酸甘油富集有益微生物群以增强香蕉抗枯萎病能力
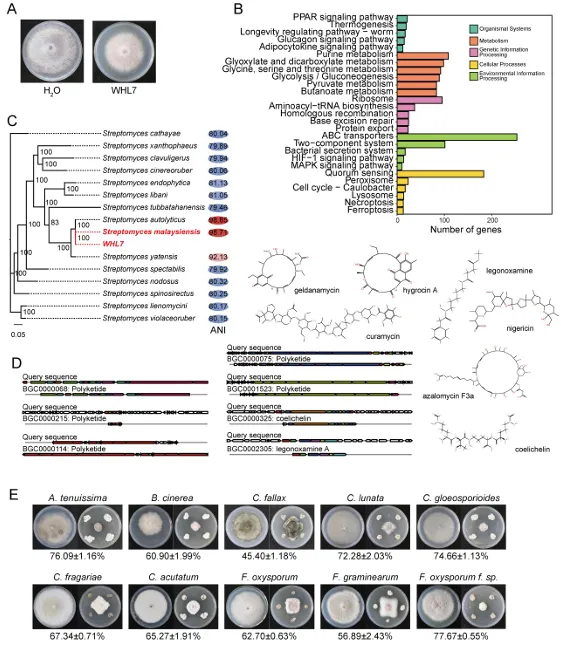
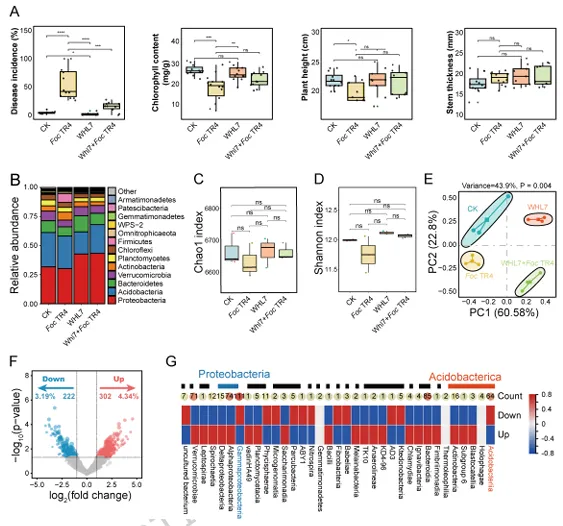
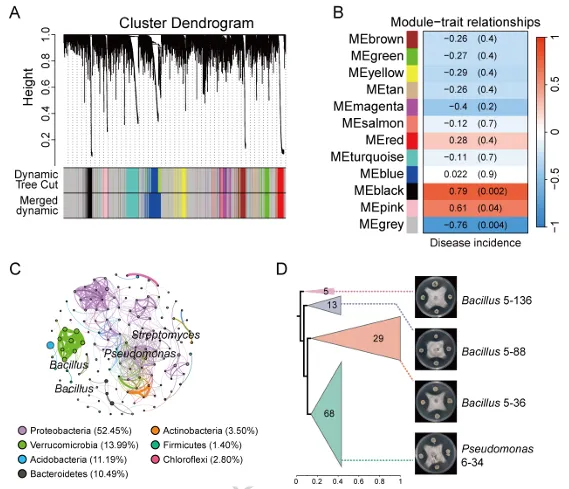
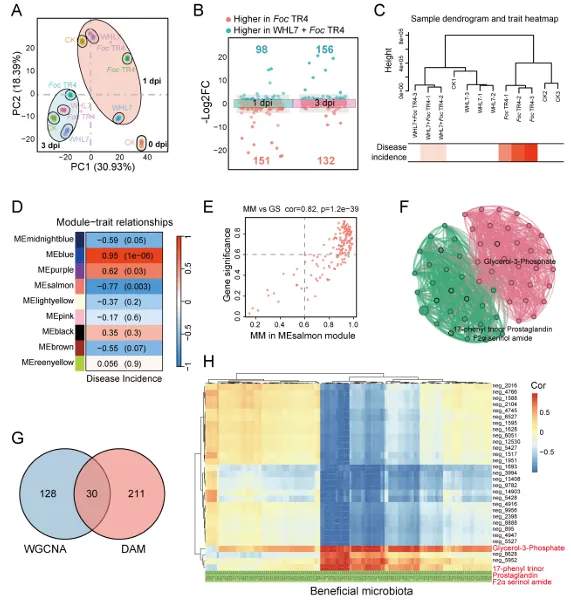
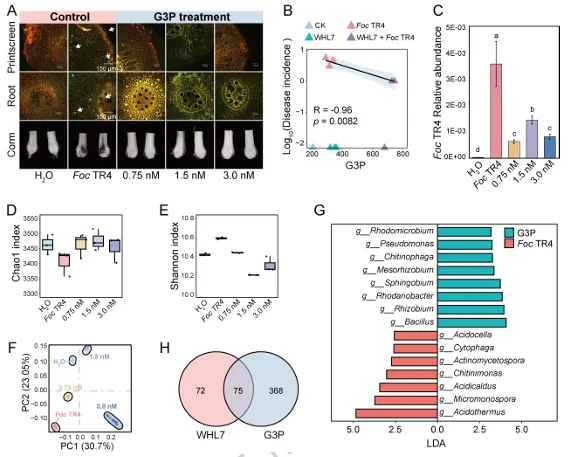
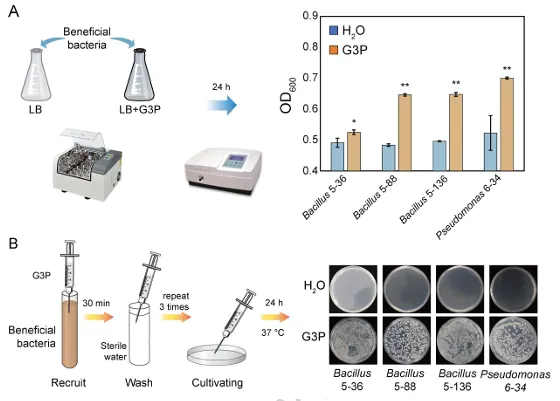
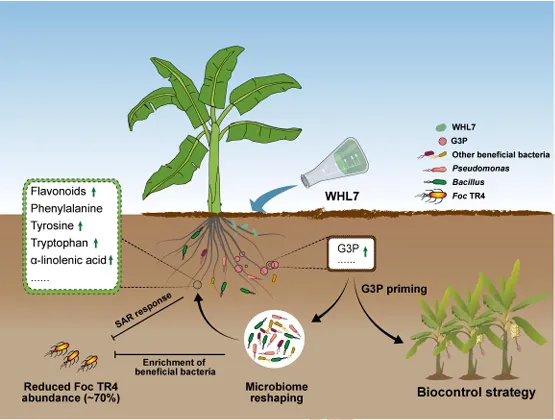
背景:尖孢镰刀菌古巴专化型热带4号生理小种(Fusarium oxysporum f. sp. cubense Tropical Race 4, Foc TR4)是香蕉枯萎病的致病菌,该病是一种破坏性土传病害。利用链霉菌(Streptomyces spp.)等拮抗微生物为真菌病害防治提供了潜在策略,但其田间应用因对微生物-植物-病原体互作机制的认知不足而受限。结果:本研究表明,海洋来源的马来西亚链霉菌WHL7(Streptomyces malaysiensis WHL7)在体外对多种植物病原真菌(尤其是Foc TR4)表现出强拮抗活性。在自然土壤盆栽实验中,WHL7处理将香蕉枯萎病发病率从Foc TR4接种对照组的55%显著降低至15%。灭菌土壤中未观察到保护效应,表明其生防效果依赖于土著微生物群落。整合代谢组学分析发现,WHL7刺激植物根系关键分泌物3-磷酸甘油(G3P)。该化合物可富集根际有益菌(如芽孢杆菌属Bacillus和假单胞菌属Pseudomonas)。与Foc TR4处理组相比,外源施加G3P使Foc TR4丰度降低73%,抑制病原侵染,并持续增加芽孢杆菌与假单胞菌的种群数量。这些富集的微生物通过诱导植物系统获得抗性(SAR)直接降低病害严重程度。结论:尽管马来西亚链霉菌WHL7在体外通过拮抗化合物直接抑制Foc TR4,但在盆栽实验中通过重塑根际微生物组间接控制病害。这种重构通过G3P介导的信号通路诱导宿主抗性。研究揭示了G3P介导的免疫启动作为土传病害可持续治理的新策略。篇名: Streptomyces-induced Glycerol-3-Phosphate enriches beneficial microbiota to enhance resistance against banana Fusarium wiltDOI: 10.1186/s40168-026-02409-6本研究提取链霉菌WHL7基因组DNA,经质量评估后利用Illumina HiSeq平台测序,并基于EggNOG数据库进行功能注释。16S rRNA基因经扩增测序及系统发育分析确定分类地位。通过双板对扣法和平板对峙法分别测定其挥发性有机物及活菌对多种植物病原真菌的拮抗活性。此外,通过盆栽实验、16S rRNA高通量测序和非靶向代谢组学分析,系统研究了WHL7对香蕉根际微生物群落结构及根系代谢谱的影响,并对关键代谢物与差异微生物进行整合网络分析。鉴定与病害相关的关键微生物类群及代谢物。进一步分离可培养有益菌,并以关键代谢物进行体外抑菌、趋化及盆栽验证,评价其抑菌效果及对根际微生物群落的调控作用。(1)海洋链霉菌WHL7具有产生多种生物活性化合物的潜力前期研究表明,WHL7提取物对Foc TR4具有强抗真菌活性。本研究进一步发现,该菌株产生的挥发性有机化合物(VOCs)同样表现出抗真菌活性,在体外能有效抑制Foc TR4的菌丝生长(图1a),表明WHL7通过代谢物与VOCs的多重拮抗策略发挥作用。我们推测其基因组可能含有负责代谢物合成的多样化生物合成基因簇(BGCs)。为此,对其基因组草图进行测序,结果显示其大小为11,820,727 bp,GC含量为69.98%,符合链霉菌属典型特征(图S1)。最长重叠群为832,367碱基,N50值为280.552 Kb(表S1)。共预测到9,310个蛋白质编码基因,其中2,813个基因通过KEGG数据库完成功能注释(表S2)。注释基因中最主要的类别为代谢相关基因(1,728个,占61.45%),其次为遗传与环境信息处理基因(424个,15.08%)和细胞过程相关基因(269个,9.57%)(图1b及表S2)。这种显著的代谢基因富集与其次级代谢产物的合成密切相关。为明确其分类地位,基于16个链霉菌基因组构建系统发育树(表S3)。对1,471个单拷贝直系同源基因的发育分析表明,WHL7与马来西亚链霉菌(Streptomyces malaysiensis)形成高支持度聚类(自举值100%,平均核苷酸一致性98.71%)(图1c,表S4)。利用antiSMASH预测WHL7基因组中的BGCs,发现其含有多组可能具抗真菌活性的聚酮类及其他次级代谢产物合成基因簇(图1d,表S5),包括格尔德霉素(相似度100%)、库拉霉素(100%)、尼日利亚菌素(94%)、潮霉素(96%)和阿扎霉素F3a(91%),同时检测到两个铁载体合成基因簇。这些丰富多样的BGCs表明WHL7具有显著的抗真菌化合物合成潜力。图1. WHL7的抗真菌活性与分类学鉴定。(A) WHL7产生的挥发性有机物(VOCs)对Foc TR4生长的抑制效应。(B) 基于KEGG数据库的WHL7基因组功能注释。(C)依据平均核苷酸一致性(ANI)值对WHL7进行物种鉴定,数据展示于末列。(D) WHL7基因组中潜在抗真菌代谢物合成基因簇预测。(E) WHL7对多种植物病原真菌的广谱抗真菌活性分析。基因组分析揭示的BGCs多样性促使我们进一步评估WHL7对10种植物病原真菌的抑制效果(图1e)。体外拮抗实验显示,WHL7具有广谱抗真菌活性,抑制率范围为45.40%至77.67%。其中对Foc TR4的抑制效果最为显著(77.67 ± 0.55%),对细极链格孢(Alternaria tenuissima,76.09 ± 1.16%)、胶孢炭疽菌(Colletotrichum gloeosporioides,74.66 ± 1.13%)和新月弯孢霉(Curvularia lunata,72.28 ± 2.03%)亦表现出强抑制活性,而对Curvularia fallax的抑制效果最弱(45.40 ± 1.18%)。这一结果证实该菌株通过多种机制介导广谱抑菌作用。(3)WHL7通过富集有益根际微生物群增强植物对Foc TR4的抗性盆栽实验表明,施用WHL7可有效抑制香蕉植株受Foc TR4侵染。在Foc TR4感染条件下,WHL7处理显著提高了叶绿素含量,但株高与茎粗无显著差异(图2a)。而在灭菌土壤中,Foc TR4单独处理组与WHL7+Foc TR4处理组间的病情严重度(图S2)及生理指标均未呈现显著差异。进一步分析显示,WHL7对Foc TR4的体外抗真菌活性与其在自然土壤中的生防效果无显著相关性(R = -0.34,p = 0.15),表明直接病原抑制并非其体内病害控制的主要机制(图S3)。WHL7的保护作用更可能通过调控根际微生物群落实现。为验证该假说,我们分析了四组处理(对照组CK、Foc TR4组F、WHL7组W、WHL7+Foc TR4组WF)的根际土壤细菌群落组成。在Foc TR4存在时,WHL7显著改变了主要细菌门的相对丰度,尤其提高了变形菌门(Proteobacteria)并降低了酸杆菌门(Acidobacteria)(图2b)。而链霉菌属(Streptomyces)的相对丰度在各处理间无显著差异(图S4),证实生防效果并非通过WHL7自身增殖直接抑制Foc TR4实现。α多样性指数(包括Chao1丰富度指数(图2c)和Shannon多样性指数(图2d))表明,WHL7缓解了Foc TR4感染导致的微生物多样性下降。β多样性分析显示四组处理的微生物群落显著分离(p = 0.004),其中WHL7+Foc TR4组与Foc TR4组样本形成独立聚类(图2e)。基于差异丰度扩增子序列变体(DAAs)的进一步分析发现,相较于Foc TR4处理组,WHL7+Foc TR4组中302个分类单元显著富集,222个分类单元显著减少(图2f)。多数DAAs属于γ-变形菌纲(Gammaproteobacteria),其中假单胞菌属(Pseudomonas)为最显著富集的菌属(图2g)。图2. WHL7处理后的生理特性与细菌群落结构。(A) 不同处理组香蕉植株的病情指数与生理指标测量。星号表示显著性差异(Student's t检验,*p<0.05,**p<0.01,***p<0.001,ns表示无显著差异)。(B) 不同样本主要细菌门的相对丰度。(C) 基于Chao1指数的细菌群落丰富度评估。(D) 基于Shannon指数的细菌群落多样性评估。(E) 主坐标分析(PCoA)展示处理组间细菌群落的β多样性。(F) Foc TR4与WHL7+Foc TR4处理组间差异ASVs分析。蓝点与红点分别代表WHL7+Foc TR4组中下调与上调的ASVs,点大小对应ASV丰度。(G) 纲水平展示两组处理间差异ASVs。(4)芽孢杆菌与假单胞菌是抑制Foc TR4侵染的核心有益菌为探究根际土壤中的核心功能菌群,我们对根际微生物群落的扩增子序列变体(ASV)丰度数据进行了加权基因共表达网络分析(WGCNA)。该分析揭示了一个与病害抑制显著相关的特定模块。WHL7处理显著减弱了微生物群落组成与发病率之间的相关性(图S5a),表明其可能通过调控微生物组结构来增强抗性。基于ASV的共表达网络被划分为12个独立模块(图3a),其中MEgrey模块被鉴定为最具生物学意义的模块。MEgrey模块与发病率的相关性分析显示两者存在显著负相关(r = -0.76,p = 0.004)(图3b),提示该模块中的微生物可能参与宿主防御反应。进一步分析发现,MEgrey模块中模块成员度(MM)与基因显著性(GS)存在极显著相关性(r = 0.58,p < 1×10^-200)(图S5b),表明网络中处于核心位置的ASVs与Foc TR4抑制密切相关,这些ASVs可能在微生物组的抗真菌功能中发挥关键作用。对MEgrey模块枢纽ASVs的分类学解析显示,变形菌门(Proteobacteria,41.22%)占比最高,其次为酸杆菌门(Acidobacteria,19.52%)和拟杆菌门(Bacteroidetes,12.41%)(图S5c)。共表达网络的详细分析鉴定出多个关键枢纽属,包括已知有益菌属芽孢杆菌(Bacillus)、假单胞菌(Pseudomonas)和链霉菌(Streptomyces),这些菌属形成了高度互连的子网络(图3c),暗示其通过协同互作维持抑病微生物组的稳定性与功能。通过整合WGCNA与差异丰度分析(DAAs),从高度保守的MEgrey模块中鉴定出143个重叠ASVs(表S6),这些ASVs均具有显著模块成员度。网络中心性与差异丰度的交汇凸显了它们的功能重要性及其与WHL7诱导的Foc TR4抑制的强关联性。为探究其功能,我们筛选了与这些枢纽ASVs对应的可培养代表菌株,并评估其体外抗Foc TR4活性。从WHL7处理香蕉植株的根际土壤中分离获得115株细菌,主要归属于芽孢杆菌属(40.87%)和假单胞菌属(59.13%)。基于系统发育树分析,从四个进化枝中各选取一株抑菌效果最佳的代表菌株进行抗Foc TR4活性分析(图3d)。PDA平板对峙实验显示,三株芽孢杆菌(5-36、5-88和5-136)和一株假单胞菌(6-34)对Foc TR4菌丝生长具有显著抑制效果(图3d),这些体外拮抗活性与其在体内抑制病害的作用相一致。综上,本研究证实WHL7通过富集根际微生物组中特定的有益类群来介导病害抑制。图3. 通过WGCNA分析鉴定病害相关ASVs。(A) 显示12个共表达ASV模块的层次聚类树。树状图每个叶片代表一个ASV,主分支展示12个模块,下方色条标注模块颜色。(B) 热图展示不同ASV共表达模块与发病率的相关性。蓝色与红色分别表示负相关与正相关,括号内数字为对应p值。(C) 采用12种方法构建的枢纽ASVs共表达网络。(D) 分离菌株系统发育树及其代表菌株对Foc TR4的抗真菌活性。不同颜色三角形代表不同进化枝,相邻数字表示各枝所含菌株数。(5)WHL7诱导的根系代谢物增强植物对Foc TR4的抗性为深入探究WHL7诱导抗性的潜在机制,我们采用非靶向代谢组学技术分析了香蕉植株在接种后0、1和3天(dpi)不同处理条件下的根系代谢谱。主成分分析(PCA)显示四组处理的代谢谱存在明显分离,其中1 dpi时的差异最为显著(图4a)。与单独接种Foc TR4或对照组相比,WHL7+Foc TR4处理的植株表现出明显的代谢重编程。比较分析鉴定出249个差异积累代谢物(DAMs),其中WHL7+Foc TR4组在1 dpi时有151个上调、98个下调,3 dpi时则有132个上调和156个下调(图4b)。42个DAMs在两个时间点均呈现一致变化(图S6a),提示其可能在持续防御调控中发挥作用。KEGG富集分析表明这些DAMs显著关联黄酮类生物合成、苯丙氨酸/酪氨酸代谢、α-亚麻酸代谢及色氨酸生物合成等关键通路(图S6b),这些通路已知参与植物防御反应、植物激素信号传导和抗菌化合物合成,表明WHL7通过调控初级代谢增强香蕉对Foc TR4的抗性。与微生物群落分析结果相似,WHL7处理显著减弱了代谢物与发病率的相关性(图4c)。通过代谢组WGCNA分析鉴定出9个共表达模块(图S7a),其中MEsalmon模块与发病率呈强负相关(r = -0.77,p = 0.003)(图4d)。该模块包含158个代谢物,WHL7处理导致其中50个显著上调、83个下调、25个无显著变化(图S7b)。值得注意的是,代谢物对发病率的重要性(GS)与模块成员度(MM)高度相关(r = 0.82,p = 1.2×10^-39),表明网络枢纽代谢物是病害抑制的核心(图4e)。通过cytoHubba插件的12种拓扑算法分析,鉴定出3-磷酸甘油(G3P)、17-苯基三烯前列腺素和F2a丝氨酸酰胺等关键枢纽代谢物(图4f)。整合WGCNA与DAMs结果发现30个代谢物在MEsalmon模块与DAMs集合中重叠(图4g和表S7)。这些枢纽代谢物与有益ASVs存在强相关性,其中G3P与多个有益细菌类群的正相关性显著强于其他代谢物(图4h)。WHL7处理还引起香蕉根系中G3P的显著积累(图S8),表明WHL7诱导G3P生物合成。因此,G3P可能作为植物-微生物互作的关键代谢物,在WHL7处理下促进有益微生物的富集。图4. 香蕉根系差异积累代谢物分析。(A) CK、WHL7、Foc TR4及WHL7+Foc TR4组在0、1和3 dpi时根系代谢物的主成分分析。(B) 火山图展示差异代谢物(DAMs)。相较于Foc TR4组,蓝点与红点分别代表1 dpi与3 dpi时WHL7+Foc TR4组中上调与下调代谢物。(C) 微生物组代表性聚类分析。(D)代谢物共表达模块与发病率相关性的热图展示。(E) MEsalmon模块中代谢物对发病率的重要性(GS)与模块成员度(MM)的散点图。(F)枢纽代谢物的共表达网络。(G) 维恩图展示MEsalmon模块中DAMs与WGCNA的重叠部分。(H) 关键代谢物与有益ASVs的整合分析。颜色表示相关系数(红:正相关,蓝:负相关)。为探究宿主来源代谢物是否介导有益根际微生物群的富集,本研究开展了外源G3P处理实验。香蕉幼苗在接种Foc TR4前分别施用不同浓度G3P,对照组仅接种Foc TR4或施用清水。通过绿色荧光蛋白(GFP)标记的Foc TR4监测病原菌定殖情况,共聚焦显微镜观察显示:接种14天后,单独Foc TR4处理的幼苗球茎和根部组织存在大量菌丝定殖,而G3P预处理显著降低病原侵染(图5a)。对四组处理(CK、Foc TR4、WHL7及WHL7+Foc TR4)的回归分析表明,G3P积累量与病害抑制呈显著负相关(R² = 0.96,p = 0.0082)(图5b)。G3P水平的升高与系统层面(而非单一处理内)的病害减轻密切相关,该相关性反映了处理方式驱动的G3P水平与病害结局变异。通过实时荧光定量PCR(qPCR)评估G3P对土壤中Foc TR4动态的影响。基于Foc TR4孢子梯度稀释建立的标准曲线(R² = 0.9906)(图S9)显示,G3P处理显著降低根际病原菌数量,其中0.75 nM浓度抑制效果最强(图5c)。体外实验证实G3P不直接抑制Foc TR4生长(图S10),表明观察到的病原定殖减少并非源于直接抗真菌活性,推测G3P通过调控根际微生物群落间接限制病原增殖。微生物组分析揭示,G3P处理导致细菌群落组成在门水平发生整体变化(图S11),与β多样性和属水平分析结果一致。α多样性分析显示,G3P部分维持了Foc TR4感染后降低的微生物丰富度(Chao1指数),但高浓度G3P(1.5 nM和3.0 nM)导致Shannon指数下降,提示群落均匀度降低可能源于特定类群的选择性富集(图5d-e)。基于主坐标分析(PCoA)的β多样性分析表明各处理组微生物群落结构显著分化,G3P处理组与单独Foc TR4组及对照组明显分离(图5f)。线性判别分析效应量(LEfSe)鉴定出G3P处理根际显著富集的细菌类群,包括WHL7处理中同样富集的芽孢杆菌属(Bacillus)和假单胞菌属(Pseudomonas),以及红微菌属(Rhodomicrobia)和中慢生根瘤菌属(Mesorhizobium)等植物促生菌(图5g)。重叠分析发现WHL7与G3P处理共同富集75个有益类群(占WHL7富集类群的52.45%),表明二者调控共同的根际微生物核心群(图5h)。由此证实,外源G3P对Foc TR4侵染的抑制效应通过重塑根际微生物群落实现。图5. G3P通过调控香蕉根际微生物群落抑制Foc TR4侵染。(A) GFP标记Foc TR4接种14天后香蕉球茎与根部的侵染症状。白色箭头指示Foc TR4绿色荧光信号,比例尺=150 μm。(B) G3P相对丰度与发病率的相关性分析,每个数据点代表独立生物学重复。(C) 实时荧光定量PCR(qRT-PCR)检测不同G3P浓度下香蕉根际土壤中Foc TR4丰度。(D) 基于Chao1指数的细菌群落丰富度评估。(E) 基于Shannon指数的细菌群落多样性评估。(F) PCoA展示不同处理组间细菌群落的β多样性。(G) G3P处理与对照组根际微生物的LEfSe分析。柱状图展示显著富集细菌类群(LDA值>2.0)的对数转换LDA得分。(H) WHL7与G3P处理富集有益微生物的重叠分析。为阐明G3P介导根际有益细菌富集的作用机制,本研究以芽孢杆菌(Bacillus)和假单胞菌(Pseudomonas)为代表菌株开展促生长与趋化性实验。生长动力学分析显示,在LB培养基中添加G3P可显著促进细菌增殖(图6a)。培养24小时后,G3P处理组的细菌在600 nm波长下的吸光度值明显高于对照组,表明G3P可作为易代谢底物支持这些细菌的生长,其中假单胞菌6-34对G3P的响应最为显著(图6a)。进一步通过趋化实验验证G3P对微生物增殖的直接刺激作用。将预处理的芽孢杆菌与假单胞菌培养24小时后(图6b),G3P处理组比清水对照组形成更密集的菌落,这与细菌对易利用碳源的趋化行为一致。类似响应也见于葡萄糖等简单糖类处理,其中芽孢杆菌5-88和假单胞菌6-34在G3P刺激下表现出更强的菌落形成能力。该结果从机制层面证实:WHL7通过诱导宿主产生G3P,进而富集芽孢杆菌与假单胞菌以抑制土传病原菌Foc TR4(图7)。图6. G3P对有益细菌的促生长与趋化作用。(A) G3P体外促进有益细菌生长速率。(B) G3P通过直接趋化作用增强微生物增殖。本研究证实WHL7在体外具有广谱抗真菌活性,尤其对Foc TR4表现出显著抑制效果。盆栽实验表明,WHL7在自然土壤中的抗真菌作用机制与平板拮抗实验存在差异。代谢组学分析揭示,WHL7接种可刺激植物根系分泌3-磷酸甘油(G3P),进而富集根际有益微生物(特别是芽孢杆菌属和假单胞菌属)。虽然G3P不直接抑制Foc TR4生长,但外源施加G3P可重现WHL7诱导的微生物群落变化并降低病原菌定殖。这些富集的有益菌通过激活关键代谢通路,诱导植物系统获得抗性。研究结果表明,WHL7通过引发G3P介导的有益菌富集间接抑制Foc TR4,而非直接拮抗作用。







 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
