3.1 GH3.8家族及匍匐翦股颖AsGH3.8的系统发育分析
为探究匍匐翦股颖中GH3.8的进化关系,利用代表性植物(包括小麦、拟南芥、多年生黑麦草和玉米)的GH3.8蛋白构建了系统发育树,发现AsGH3.8与小麦TaGH3.8同源性最高。进一步的蛋白序列比对表明,这些GH3.8成员均具有一个高度保守的Pfam结构域,这是该蛋白家族特有的功能区域(Fig. 1)。
图1. AsGH3.8的系统发育分析。 (a) 基于邻接法使用MEGA7构建的不同物种系统发育树。(b) 氨基酸序列比对。红框表示GH3.8蛋白特有的Pfam功能结构域。
3.2. 过表达AsGH3.8匍匐翦股颖的表型特征
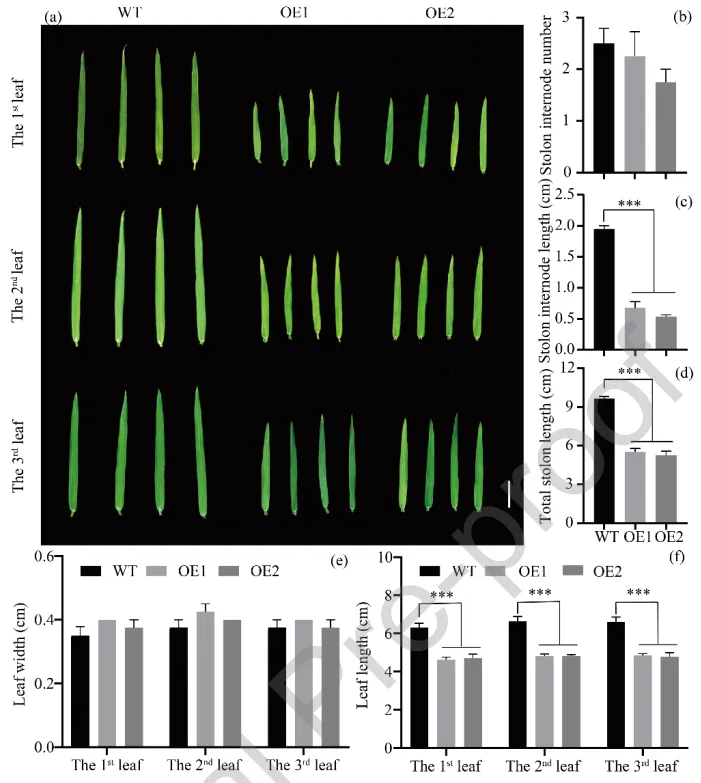
为探究AsGH3.8在植物生长中的潜在作用,在匍匐翦股颖中过表达该基因并筛选出高表达株系(OE1和OE2)。表型观察显示,过表达AsGH3.8显著抑制了植株生长,转基因株系的完全展开叶长度、匍匐茎节间长度及匍匐茎总长度均显著低于野生型,但叶宽和匍匐茎节间数量无显著差异(Fig. 2)。
图2. 过表达AsGH3.8匍匐翦股颖的表型特征。 (a) 匍匐翦股颖株系的叶片表型。比例尺 = 1 cm。(b) 匍匐茎节间数量。(c) 匍匐茎节间长度。(d) 匍匐茎总长度。(e) 叶宽。(f) 叶长。表示根据学生t检验,WT与AsGH3.8*-OE株系之间存在显著差异。显著性水平:*** p < 0.001。误差棒代表标准误(n = 5)。
3.3. 升高[CO₂]下过表达AsGH3.8匍匐翦股颖的表型特征
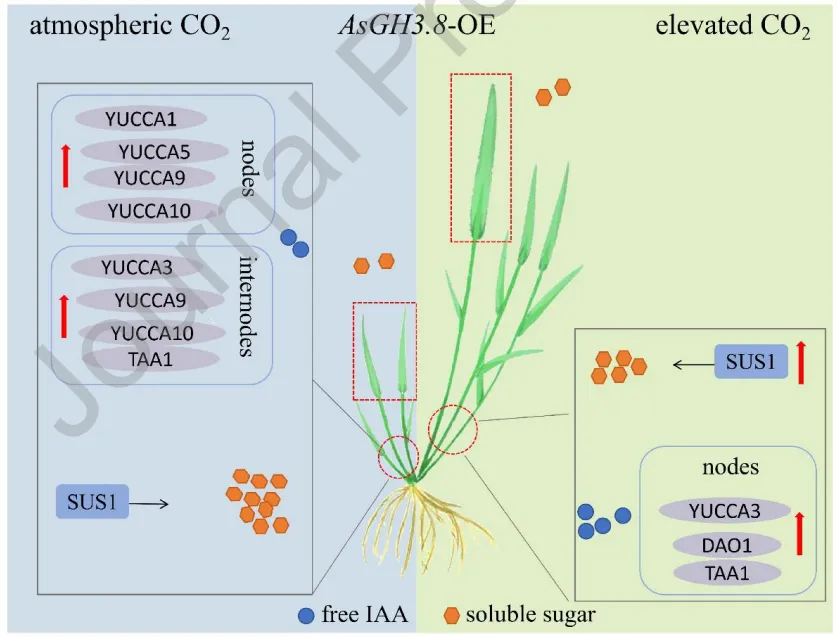
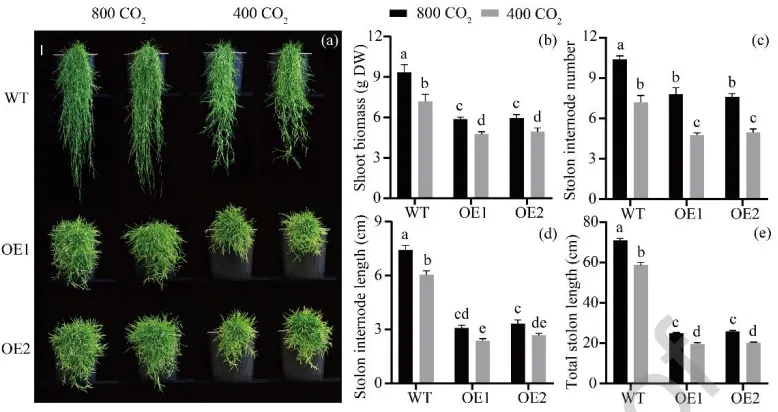
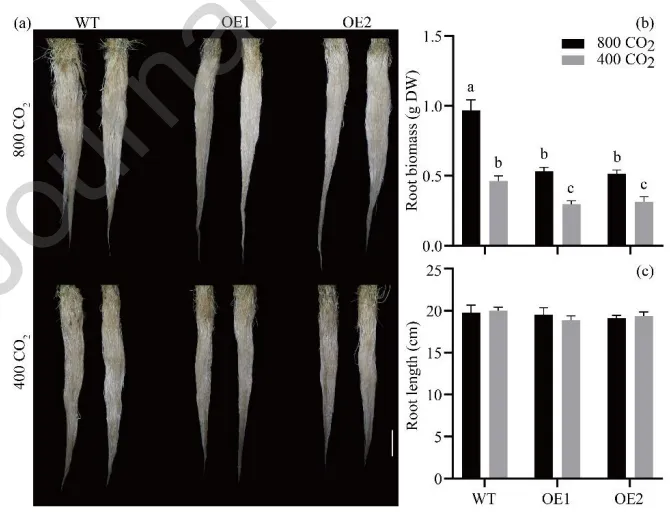
在环境和升高[CO₂]条件下,评估了AsGH3.8基因及[CO₂]对植物生长的影响。结果表明,无论在何种[CO₂]水平下,过表达株系的地上部和根系生物量、匍匐茎节间数量及长度等指标均显著低于野生型。然而,与环境[CO₂]相比,升高[CO₂]显著促进了野生型和转基因株系的地上部和根系生物量积累以及匍匐茎的伸长与节间发育,但对根长无显著影响(Fig. 3,Fig. 4)。
图3. [CO₂]对过表达AsGH3.8和WT匍匐翦股颖地上部表型的影响。 比例尺 = 5 cm。(a) 地上部表型。(b) 地上部生物量。(c) 匍匐茎节间数量。(d) 匍匐茎节间长度。(e) 匍匐茎总长度。在每个图中,不同字母表示根据F检验(p < 0.05),条形图中各处理比较存在显著差异。误差棒代表标准误(n = 5)。DW = 干重。
图4. [CO₂]对过表达AsGH3.8和WT匍匐翦股颖根系表型的影响。 比例尺 = 3 cm。(a) 根系表型。(b) 根系生物量。(c) 根长。在每个图中,不同字母表示根据F检验(p < 0.05),条形图中各处理比较存在显著差异。误差棒代表标准误(n = 5)。DW = 干重。
3.4. 过表达AsGH3.8匍匐翦股颖内源生长素含量分析
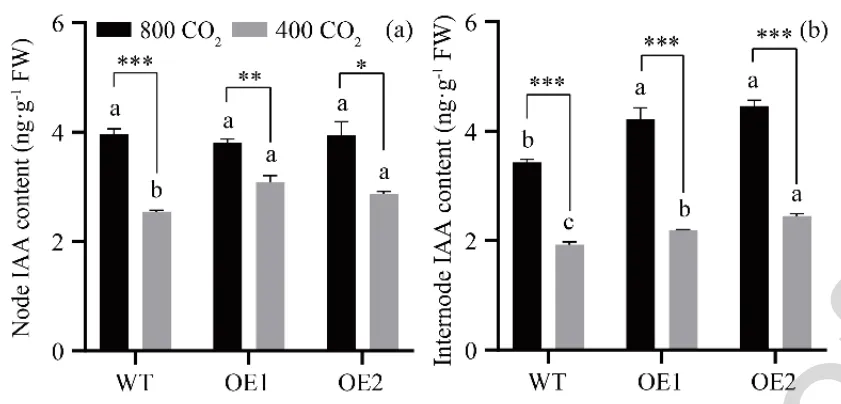
为探究AsGH3.8在升高[CO₂]下调控植物生长的潜在作用,对内源IAA含量进行了定量分析。在环境[CO₂]下,过表达株系节和节间的IAA含量均高于野生型。在升高[CO₂]下,过表达株系节间的IAA含量显著高于野生型,但节中无显著差异。与环境[CO₂]相比,升高[CO₂]显著提升了野生型和过表达株系节与节间中的IAA水平,大幅促进了匍匐茎中IAA的积累(Fig. 5)。
图5. [CO₂]对过表达AsGH3.8和WT匍匐翦股颖节和节间IAA含量的影响。 (a) 节IAA含量。(b) 节间IAA含量。在每个图中,不同字母分别表示根据F检验(p < 0.05),在环境或升高[CO₂]下WT与AsGH3.8-OE株系之间存在显著差异。表示根据学生t检验,环境和升高[CO₂]之间存在显著差异。显著性水平:p < 0.05,** p < 0.01,*** p < 0.001。误差棒代表标准误(n = 3)。FW = 鲜重。
3.5. 过表达AsGH3.8匍匐翦股颖中生长素代谢相关基因的表达
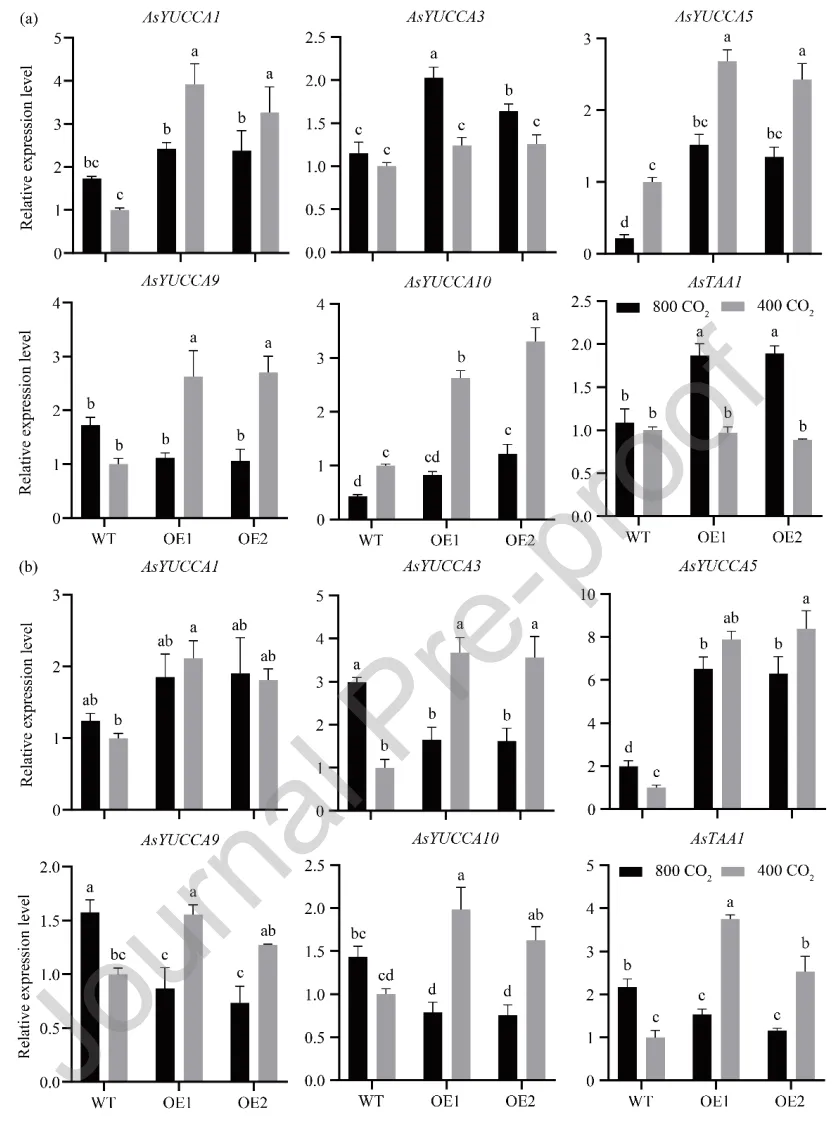
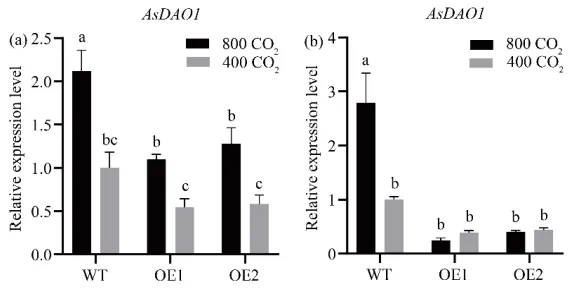
为确定升高[CO₂]对生长素代谢基因表达的影响,分析了节和节间中生长素合成(AsYUCCA系列、AsTAA1)及氧化(AsDAO1)基因的表达水平。在节中,环境[CO₂]下过表达株系多个合成基因显著上调;升高[CO₂]诱导过表达株系部分合成基因上调,同时下调了氧化基因AsDAO1。在节间中,环境[CO₂]下过表达株系多数合成基因上调;而升高[CO₂]则导致过表达株系中除AsYUCCA5外的多数合成基因及氧化基因表达下调。这表明升高[CO₂]对野生型和转基因株系的生长素代谢基因引发了截然不同的转录重编程(Fig. 6,Fig. 7)。
图6. [CO₂]对过表达AsGH3.8和WT匍匐翦股颖节和节间生长素合成基因表达水平的影响。 (a) 节中的基因表达。(b) 节间中的基因表达。在每个图中,不同字母表示根据F检验(p < 0.05),条形图中各处理比较存在显著差异。误差棒代表标准误(n = 3)。
图7. [CO₂]对过表达AsGH3.8和WT匍匐翦股颖节和节间生长素氧化基因表达水平的影响。 (a) 节中AsDAO1的表达。(b) 节间中AsDAO1的表达。在每个图中,不同字母表示根据F检验(p < 0.05),条形图中各处理比较存在显著差异。误差棒代表标准误(n = 3)。
3.6. 过表达AsGH3.8匍匐翦股颖的可溶性糖含量
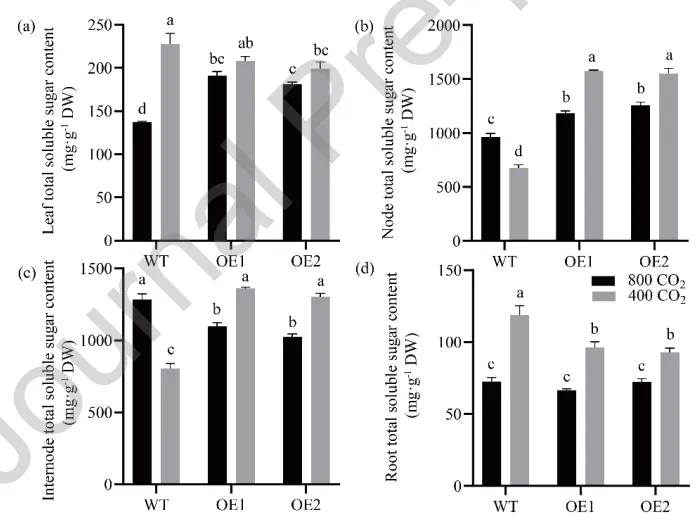
测定了不同组织中的总可溶性糖含量以评估碳分配。在叶片中,升高[CO₂]显著降低了野生型的糖含量,而过表达株系维持较高水平。在匍匐茎节和节间中,环境[CO₂]下过表达株系的糖含量均显著高于野生型;当处于升高[CO₂]时,野生型的糖含量显著增加,而过表达株系的糖含量则出现显著下降。在根系中,升高[CO₂]同样导致野生型和过表达株系的总可溶性糖含量显著降低(Fig. 8)。
图8. [CO₂]对过表达AsGH3.8和WT匍匐翦股颖叶片、节、节间和根系中总可溶性糖含量的影响。 (a) 叶片总可溶性糖含量。(b) 节总可溶性糖含量。(c) 节间总可溶性糖含量。(d) 根系总可溶性糖含量。在每个图中,不同字母表示根据F检验(p < 0.05),条形图中各处理比较存在显著差异。误差棒代表标准误(n = 4)。DW = 干重。
3.7. 过表达AsGH3.8匍匐翦股颖中蔗糖合成基因的表达
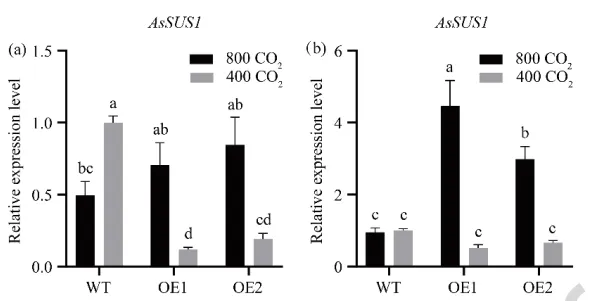
分析了蔗糖合成基因AsSUS1的表达模式。在匍匐茎节中,环境[CO₂]下过表达株系的AsSUS1表达量显著低于野生型;但在升高[CO₂]下,野生型表达下调,而过表达株系表达上调。在匍匐茎节间中,环境[CO₂]下两株系无显著差异;而在升高[CO₂]处理下,过表达株系中AsSUS1的表达量被显著诱导并高于野生型,表明升高[CO₂]激活了转基因株系中蔗糖代谢相关基因的转录(Fig. 9)。
图9. [CO₂]对过表达AsGH3.8和WT匍匐翦股颖节和节间蔗糖合成基因表达水平的影响。 (a) 节中AsSUS1的表达。(b) 节间中AsSUS1的表达。在每个图中,不同字母表示根据F检验(p < 0.05),条形图中各处理比较存在显著差异。误差棒代表标准误(n = 3)。








 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
