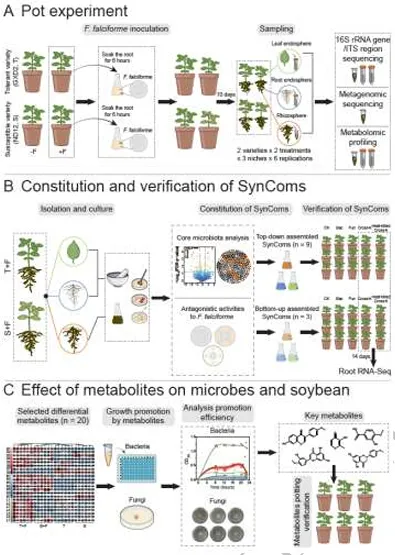
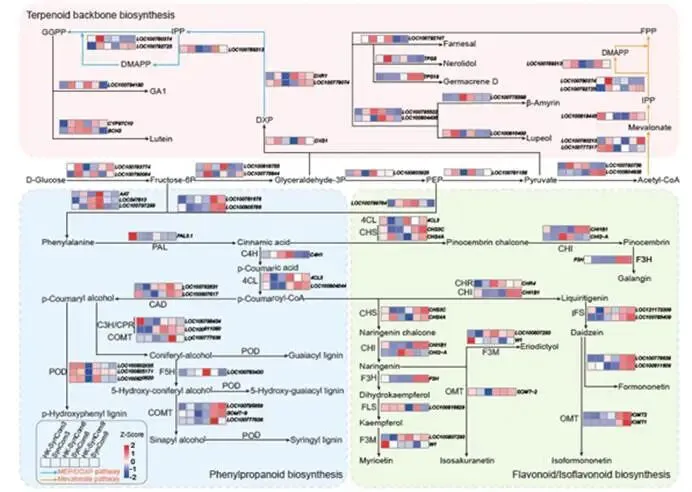
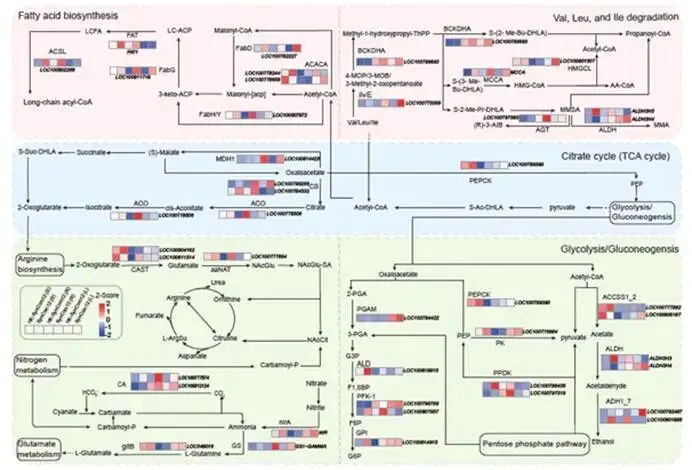
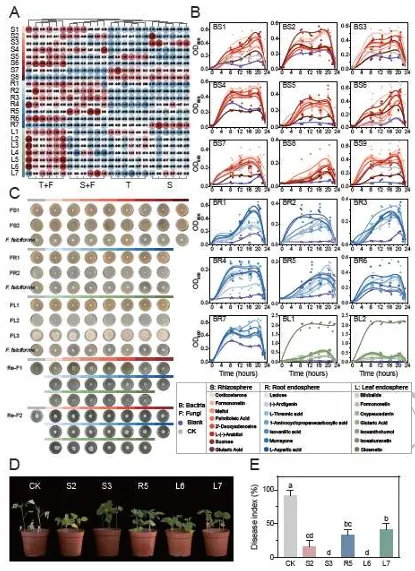
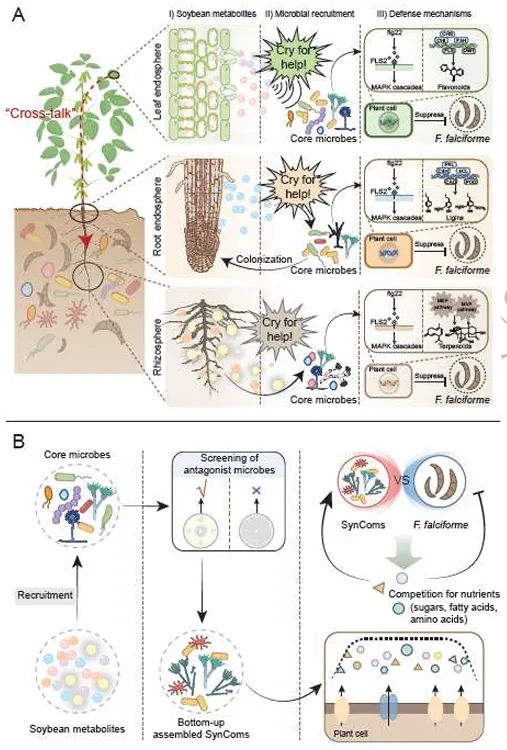
ISME Journal | 华南农业大学:跨生态位代谢物-微生物组互作协同调控大豆对镰刀菌根腐病的系统抗性
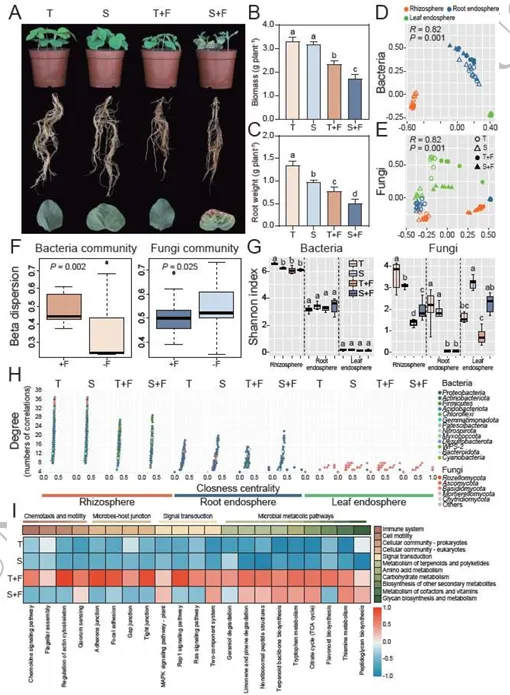
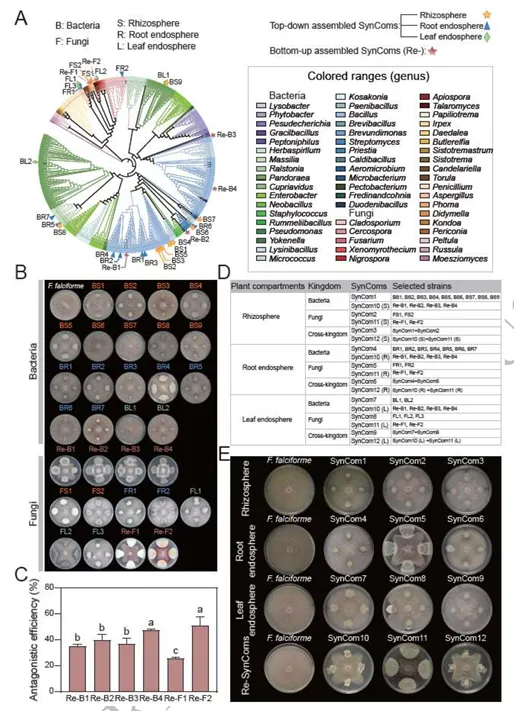
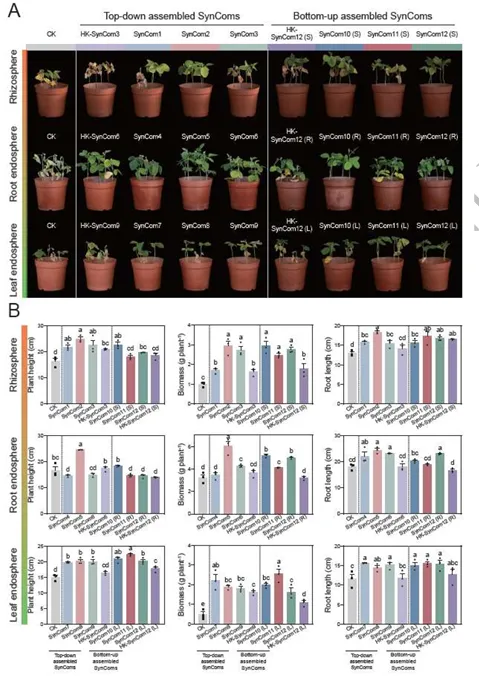
由镰刀菌(Fusarium falciforme)主导引发的大豆根腐病(Fusarium root rot)对全球大豆生产构成严重威胁。基于微生物组的策略虽能提供可持续防治方案,但多生态位互作的机制尚不明确。本研究揭示,耐病大豆品种(GXD2)通过协调空间分辨的代谢物信号,在根际、根内圈和叶内圈系统性招募有益微生物:其中,芒柄花素(formononetin)和麦芽酚(maltol)选择性富集根际中的芽孢杆菌(Bacillus)和马赛菌(Massilia);牛蒡苷元(arctigenin)和异香草酸(isovanillic acid)招募根内圈的芽孢杆菌与链霉菌(Streptomyces);而黄酮类化合物(如香叶木素diosmetin)则吸引叶内圈的青霉菌(Penicillium)和曲霉菌(Aspergillus)。基于这些互作机制,我们通过自上而下(宿主选择类群)和自下而上(基于拮抗剂)策略构建了不同类型的合成微生物群落(SynComs)。两种SynComs均能抑制感病品种的根腐病,其中叶面喷施自上而下型SynComs可显著促进地上部生长。转录组分析表明,二者作用模式存在差异:自上而下型SynComs激活了丝裂原活化蛋白激酶(MAPK)关联的萜类与黄酮代谢通路,而自下而上型SynComs主要调控宿主碳氮分配,从而有效限制病原体资源获取。本研究首次提出“代谢物介导的多生态位协同防御”模型,为微生物组驱动的病害治理提供了理论框架,并为可持续作物保护策略开辟了新路径。篇名: Cross-niche metabolite-microbiome interactions orchestrate systemic soybean resistance to Fusarium root rotDOI: 10.1093/ismejo/wrag080(A) 盆栽实验设计:采用两种基因型大豆(耐病型T/感病型S),分别进行镰刀菌接种与模拟处理。接种10天后采集根际、根内圈及叶内圈样本(每组n=6)。通过16S rRNA基因和ITS区扩增子测序分析群落组成,宏基因组测序评估根际微生物功能,代谢组学分析与代谢物检测阐明大豆-微生物互作机制。(B) 合成菌群构建与功能验证:从接种镰刀菌的健康与发病植株根际、根内圈及叶内圈分离微生物菌株。整合富集微生物分析(自上而下策略)与体外镰刀菌拮抗实验(自下而上策略)结果,组装12种SynComs并接种至感病品种筛选最优组合,通过转录组测序(RNA-seq)鉴定差异表达基因。(C) 代谢物对微生物及大豆的影响:从根际、根内圈和叶内圈筛选20种代谢物,测试其对细菌/真菌菌株的促生长能力,分析代谢物-菌株互作关系,并通过盆栽实验评估其病害缓解效应。(1)镰刀菌接种引发严重根腐症状及基因型依赖的定殖为探究不同基因型大豆品种对镰刀菌侵染的响应,本研究结合多组学方法及多种验证手段开展了一系列实验(图1)。盆栽试验中,接种镰刀菌的植株表现出高发病率,并呈现典型的根腐病症状:地上部生长受阻、根系褐变腐烂、叶片黄化或萎蔫,部分植株死亡(图2A)。相应地,耐病品种(GXD2)的生物量在接种后由3.32 g降至2.35 g,而感病品种(ND12)的降幅更为显著,其生物量从对照组的3.19 g锐减至处理组的1.73 g(方差分析,P < 0.01,n = 4;图2B-C)。为验证症状严重程度与病原菌负载量的相关性,我们定量分析了接种后0、5和10天(dpi)根际中镰刀菌的拷贝数(标准曲线与熔解曲线见补充材料图S1)。尽管两品种接种后病原菌负载量均增加,但耐病品种(GXD2)在5 dpi后表现出显著抑制,至10 dpi时其病原菌丰度仍比持续激增的感病品种低5.53倍(图S2A)。这些结果表明,镰刀菌侵染严重抑制大豆生长,且根系定殖程度及病害严重性受宿主基因型的强烈调控。我们表征了接种与非接种条件下两个大豆品种根际、根内圈及叶内圈的细菌与真菌群落(图1A)。测序共获得72个样本的4,340,452条高质量16S rRNA基因序列和5,591,052条ITS1序列,经去冗余后基于精确序列分辨率得到6,874个细菌ASV(扩增子序列变体)和2,917个真菌ASV。β最近分类单元指数(βNTI)分析表明,镰刀菌侵染后微生物群落发生适度重构(双尾学生t检验,P < 0.05)。接种处理导致除根内圈微生物组外,所有区室的细菌与真菌群落的βNTI值均发生偏移(图S2B、S2C)。在根内圈,感病品种的同质性选择比例(βNTI < -2,93.3%)显著高于耐病品种(73.3%),表明感病植株在感染后受到更严格的环境过滤(图S2D、S2F)。跨区室分析显示,真菌的确定性选择(βNTI < -2)在根际和叶内圈增强,而接种处理降低了真菌的确定性信号,却增强了根内细菌的确定性选择(图S2F、S2G)。这些结果说明耐病品种根内圈的细菌群落经镰刀菌侵染后发生更严格的确定性重组,而真菌组分相对不受宿主基因型影响。PERMANOVA分析证实空间生态位是微生物组结构的主要驱动因素,区室效应分别解释了细菌组成74.0%和真菌组成26.0%的变异(均P < 0.001;表S4)。相比之下,接种状态与宿主基因型解释的变异比例较小但仍具统计学意义。与此一致,Bray-Curtis主坐标分析(PCoA)沿PC1轴清晰分离了各区室的细菌群落,并将真菌群落划分为地下与地上集群(图2D-E、图S3)。尽管门水平细菌组成在区室间呈现明显分类学差异,真菌群落则表现出更保守的门水平分布(尽管整体结构存在显著差异)(图S4)。β离散度分析表明接种后细菌群落的异质性增强(图2F、表S5)。根内圈中,耐病与感病品种的细菌组成差异显著,而真菌群落无显著变化(表S6)。线性混合模型分析进一步强化上述趋势。细菌α多样性主要受区室影响,而真菌α多样性的决定因素则依赖于度量指标:镰刀菌接种是真菌Shannon指数的首要驱动因子,而区室对Observed ASVs丰富度的影响更强(均P < 0.001;表S7)。Shannon指数从根际到根内圈递减,且病原菌挑战降低了根际与根内圈真菌多样性,但叶内圈未出现显著下降(图2G)。基因型效应对真菌Shannon指数(P = 0.005)和细菌Observed ASVs丰富度(P = 0.017)虽较弱但仍显著。综上,空间生态位是大豆微生物组的主要生态过滤器,但病原菌入侵叠加了基因型依赖的重构效应,这一现象在耐病品种根内圈的细菌群落中最为显著。图2 耐病(T)与感病(S)大豆在镰刀菌胁迫下的表型与多组学特征。(A) 接种后地上部、根系及叶片表型图。(B-C) 全株及根系鲜重(均值±SD,n=4)。(D-E) 根际、根内圈与叶内圈中细菌(D)和真菌(E)群落的主坐标分析(PCoA)。(F)接种(+F)与未接种(-F)条件下微生物群落的β-离散度。(G) 三区室中细菌/真菌群落的香农指数。(H) 关联度数与接近中心性的网络拓扑特征。(I) 趋化性、信号转导及代谢通路的宏基因组功能热图(色标示标准化丰度:红色高富集,蓝色低丰度)。不同小写字母表示统计学差异(单因素ANOVA,P < 0.05)。共现网络在各区室间差异显著,且镰刀菌侵染以基因型依赖的方式重塑网络结构(图S5,表S8)。在根际与根内圈网络中,侵染通常导致网络规模缩小(节点/边减少),而叶内圈网络始终稀疏。侵染条件下(+F),耐病基因型根内圈网络比感病基因型更小且连接度更低(图2H,表S8),这与宿主过滤更严格的群落特征一致,可能有助于抵抗镰刀菌。根际网络中,两基因型接种后负连接比例均下降,但感病品种(ND12)始终高于耐病品种(GXD2)(表S8),表明ND12在病原压力下具有相对更具竞争性的关联结构。枢纽分析(介数中心度前1%)发现,根际网络中的枢纽类群包括芽孢杆菌(Bacillus)、链霉菌(Streptomyces)、青霉菌(Penicillium)和一个未分类曲霉科(Aspergillaceae)ASV;根内圈网络以马赛菌(Massilia)为枢纽;叶内圈网络则以枝孢菌(Cladosporium)为枢纽(表S9)。差异丰度分析显示接种后存在强烈的区室特异性招募(图S6)。感病品种中,根际、根内圈和叶内圈分别富集了92、27和0个细菌ASV及23、1和1个真菌ASV;耐病品种则在各生态位富集81、36和0个细菌ASV,且每个生态位仅1个真菌ASV,表明其真菌群落变化有限。跨区室汇总显示,感病植株新增119个细菌和25个真菌ASV,而耐病植株新增117个细菌但仅3个真菌ASV(表S10)。直接基因型对比发现,耐病品种根际富集22个细菌和2个真菌ASV,根内圈富集28个细菌ASV,叶内圈富集2个真菌ASV(表S11)。响应最显著的类群包括固氮螺菌(Azospirillum)、芽孢杆菌、类芽孢杆菌(Paenibacillus)、短芽孢杆菌(Brevibacillus)、马赛菌和青霉菌(图S7A)。耐病植株还累积了曲霉(Aspergillus)、青霉和黑孢霉(Nigrospora)ASV(图S7B–C),且根际显著富集潘多拉菌(Pandoraea),根内圈富集草螺菌(Herbaspirillum)(图S7D–F)。综上,耐病大豆品种优先招募潜在有益细菌类群,而几乎排斥额外真菌响应者。宏基因组分析揭示了镰刀菌侵染后根际微生物组发生显著的基因型依赖性功能转变。经质量控制后,宏基因组共包含4,987个细菌和442个真菌分类单元(均带基因注释)。基于KEGG直系同源(KO)、COG、CAZy和MetaCyc谱的非度量多维标度分析显示,接种与非接种样本沿MDS1轴明显分离(PERMANOVA,多数情况下R > 0.67,P = 0.001;图S8A),而植物基因型则使样本沿MDS2轴进一步分化。差异丰度分析表明,接种显著减少了两基因型中KO、COG和MetaCyc特征的数量,但CAZyome规模未发生明显变化(图S8B)。尽管整体功能库收缩,耐病品种根际仍保留了若干过度富集的功能类别。与感病植株相比,耐病植株中与趋化性、鞭毛组装、信号转导及专一性代谢相关的基因显著更丰富(图2I)。典型实例包括:鞭毛马达开关基因fliG(K02410)、色氨酸代谢单加氧酶基因(K03392)以及参与类黄酮降解的香叶基香叶基转移酶基因(K01692),这些基因在接种后耐病微生物组中的丰度比感病组高29.2%-59.4%(单因素方差分析,P < 0.05;图S8C-E)。次级代谢物生物合成通路的功能富集主要归因于耐病根际中链霉菌、假单胞菌和芽孢杆菌的相对丰度更高(图S6F)。这些结果表明,尽管镰刀菌广泛缩减了富集功能库,但耐病基因型仍保留甚至放大了与运动性、信号传导及抗菌代谢相关的关键功能特征。(5)合成微生物群落(SynCom)促进感病大豆生长并抑制根腐症状通过梯度稀释平板法,从耐病与感病大豆的根际、根内圈及叶内圈共获得476株可培养菌株(图3A,NCBI生物项目PRJNA1433828)。经去冗余和Sanger测序鉴定,共包含50种独特细菌和39种独特真菌。双培养实验将10株菌归类为接触依赖性拮抗菌,9株为扩散性抗生素产生菌,44株为非拮抗菌(图3B,图S9)。拮抗菌以芽孢杆菌、嗜热芽孢杆菌、篮状菌、枝孢菌和青霉菌为主(图3C,表S10)。根据两种设计原则构建了12种SynCom:第一种基于耐病品种选择性富集的类群(自上而下策略),第二种包含对镰刀菌具强拮抗性的菌株(自下而上策略)及其组合(图3D-E,表S12)。通过生长动力学和缺失实验验证了SynCom的稳定性(图S10-S14)。完整SynCom在48小时后的饱和密度(OD600)显著高于单一菌株(P < 0.05;图S10E-H),表明存在生态位互补性。缺失实验发现芽孢杆菌BR1为关键结构供体,其移除导致群落生物量显著降低(图S12G)。图3 合成菌群(SynComs)构建及其对镰刀菌的抑制能力。(A) 从根际、根内圈与叶内圈分离的可培养细菌/真菌的最大似然系统发育树(彩色分支表示SynComs组装所选类群)。(B) 双培养平板显示各生态位代表性菌株对镰刀菌的生长抑制。(C) 四种高活性细菌与两种高活性真菌的抑制率(均值±SD,n=3;不同字母示显著差异,单因素ANOVA,P < 0.05)。(D) 12种跨界SynComs设计:SynComs 1-9为区室特异性;SynComs 10-12通过土壤浇灌(根际S)、根系浸蘸(根内圈R)或叶面喷雾(叶内圈L)施用于全区室。(E) 12种SynComs对镰刀菌的径向生长抑制效果比较。在感病品种ND12中,所有活性SynCom均较水对照(CK)减轻病害(图4A)。跨界组合(Cross-K)在所有实验中均表现稳健且一致的病害抑制效果(图4B,图S15)。但真菌SynCom在特定区室(尤其是根内圈或叶内圈)表现出更显著的促生长作用(图4B)。基于耐病富集类群的SynCom在根和叶内圈的病害指数更低(方差分析;图S15B-C)。出乎意料的是,热灭活根内圈SynCom(HK-Cross-K)仍能提供部分保护,提示存在代谢物介导的效应。定殖实验显示群落的持久性具有选择性:目标真菌和马赛菌BR5成功占据优势生态位(图S16-S17),而草螺菌BR7未能持续存在。这些数据表明SynCom通过关键菌驱动的生物量稳定性和宿主选择的生态位占据共同建立防御屏障。对6种跨界SynCom(3种耐病富集型:SynCom 3/6/9;3种拮抗剂型:均基于SynCom 12)进行转录组分析(图4A-B,表S13)。主成分分析(PCA)显示SynCom处理组与对照组沿PC1轴(72.3%方差)分离(图S9A-B)。严格筛选(|log2FC| ≥ 1,FDR < 0.05)发现,根中自上而下型SynCom(源自根际、根内圈和叶内圈类群)分别上调605、435和243个差异表达基因(DEG)(图S18A-B);自下而上型SynCom诱导更大转录变化(上调2,377/4,249/527个DEG),表明免疫激活具有策略依赖性(图S18C)。图4 合成菌群(SynComs)介导的感病大豆镰刀菌根腐病缓解效应。(A) ND12品种接种不同SynComs 14天后的表型。(B) 株高、地上部生物量及根长(均值±SD,n=3;相同字母示无显著差异,单因素ANOVA,P < 0.05)。KEGG富集分析揭示双重防御机制:自上而下型跨界SynCom在根际上调萜类骨架合成(ko00900),在根内圈上调苯丙烷合成(ko00940),在叶内圈上调黄酮/异黄酮合成(ko00941)(图5,图S18D);而拮抗剂型SynCom主要重塑初级代谢,增强氮代谢(ko00910)、生物素代谢(ko00780)和糖酵解/糖异生(ko00010)(图6,图S18D)。图5 萜类骨架、苯丙烷及黄酮/异黄酮生物合成通路的差异表达图谱。热图显示活体跨界SynComs(根际来源SynCom3、根内圈来源SynCom6、叶内圈来源SynCom9)与其热灭活对照(HK-SynCom3/6/9)的转录水平差异。色阶表示相对于热灭活对照的log2倍数变化(蓝色:下调,红色:上调)。图6 拮抗型SynComs接种后核心初级代谢通路的差异表达。热图展示三羧酸循环(TCA)、糖酵解/糖异生、脂肪酸合成及氮代谢途径中关键酶的转录水平。比较基于镰刀菌拮抗菌株构建的活体跨界SynComs(根际SynCom12(S)、根内圈SynCom12(R)、叶内圈SynCom12(L))与其热灭活对照(HK-SynCom12(S/R/L))。色阶表示相对于配对热灭活对照的log2倍数变化(红色:上调,蓝色:下调);白色示该比较中未检测到基因表达。综上,生理与转录组数据表明,合理设计的SynCom能挽救感病大豆免受镰刀菌根腐病侵害,其中跨界组合通过协同激活系统抗性(ISR)、强化专一性代谢通路及重构中心代谢,提供最强保护效果。(6)镰刀菌重塑区室代谢组并调控微生物-代谢物互作非靶向LC-MS分析表明,病原菌侵染后根系分泌物的化学组成发生了显著的区室特异性重塑。OPLS-DA模型显示,在两个品种的根际(R²X=0.68,Q²=0.95)、根内圈(R²X=0.75,Q²=0.95)和叶内圈(R²X=0.57,Q²=0.97)中,接种组与对照组样本均呈现明显分离(置换检验,P<0.01;图S19A)。通过火山图和维恩图分析,我们在各区室中鉴定出了差异丰度代谢物(|log2FC|≥1,P<0.05)(图S19B)。具体而言:在根际区域,耐病品种显著积累了芒柄花素(formononetin,log2FC=1.10,P<0.05)和麦芽酚(maltol,log2FC=1.25,P<0.01),而两个基因型共同表现出皮质酮、棕榈油酸和L-阿拉伯糖醇含量的增加(图7A)。在根内圈区域,241种富集代谢物中,耐病品种的苯丙烷类中间体(如牛蒡苷元arctigenin、异香草酸isovanillic acid和九里香酮murrayone)显著上调(图7A)。在叶内圈区域,次生代谢物普遍增加,耐病品种特异性富集了黄酮类和萜类化合物,如白果内酯(bilobalide)、氧化前胡素(oxypeucedanin)、异黄腐醇(isoxanthohumol)、异樱花素(isosakuranetin)和香叶木素(diosmetin)(图7A)。我们选取了20种指示性化合物(每个区室排名前7-8位,包括2种共有代谢物)进行镰刀菌和代表性SynCom菌株的互作测试(图7B-C,图S20A)。结果显示,大多数化合物主要通过促进有益菌生长而非直接抑制病原菌来发挥作用(图7C)。这种促生长效应具有高度底物特异性,例如马赛菌能够利用芒柄花素(OD600增加3.2-14.9%)和棕榈油酸(OD600增加25.9-90.4%),但会被其他代谢物抑制(图7B)。耐病基因型富集的代谢物不仅促进了芽孢杆菌、链霉菌、类芽孢杆菌和草螺菌等细菌的生长,还对真菌内生菌产生了显著的正向影响(图7B-C)。通过ImageJ定量分析发现,这些抗性相关代谢物显著增加了青霉菌和曲霉菌的菌落面积,但对黑孢霉的影响较小(图S21)。这些结果表明,抗性相关代谢物通过选择性促进有益细菌和真菌类群的增殖,重塑了宿主-微生物的互作网络。图7 靶标代谢物促进有益微生物并抑制镰刀菌根腐病。(A) 气泡图显示接种(+F)与未接种(-F)条件下耐病(GXD2)与感病(ND12)大豆各区室关键代谢物的富集情况(气泡大小示倍数变化,颜色示显著性,t-检验P < 0.05)。(B) OD600测定(24小时)显示选定代谢物对各区室SynCom细菌成员的生长影响(相对溶剂对照的百分比)。(C) 对SynCom真菌及镰刀菌的相应效应。(D) 五种代谢物单独处理(按原生区室采用土壤浇灌或叶面喷施)的盆栽试验表型。(E) 代谢物处理组的病情指数降低(均值±SD,n=6;不同字母示显著差异,单因素ANOVA,P < 0.05)。为验证体外促进有益菌生长的代谢物是否能在植物体内赋予抗病性,我们选取了五种关键代谢物(芒柄花素、麦芽酚、异香草酸、异樱花素和香叶木素)研究其对大豆根腐病的调控作用。盆栽实验显示,外源添加这些代谢物使大豆根长增加14.2%-41.5%,叶绿素含量(SPAD值)提高2.86-3.36倍,生物量积累增加43.6%-206.3%(图S20B-E)。代谢物处理组的根腐病病情指数显著低于对照组(P<0.01;图7D-E)。这些结果系统揭示了植物通过代谢组重塑精确调控微生物群落,从而形成抵抗土传病害的"代谢屏障"的生态适应机制。本研究提出并验证了一种"代谢物-微生物组耦合的多生态位协同防御"模型。耐病大豆品种通过特异性代谢产物精确招募跨界有益微生物组,在双向地上部-根系信号调控下构建全株免疫屏障。通过自上而下(宿主选择核心类群)和自下而上(拮抗菌株筛选)策略组装的合成菌群(SynComs)均显著减轻病害症状,但其分子与功能机制存在差异(图8)。特别值得注意的是,研究数据揭示了常被忽视的真菌伙伴的重要贡献:将青霉菌(Penicillium)、黑孢霉(Nigrospora)及相关菌株纳入SynComs后,其植株保护效果优于纯细菌组合,更能真实还原天然的跨界微生物组复杂性。这些发现深化了我们对植物-微生物组协同防御根腐病的理解,为跨界SynComs的理性设计提供了可靠框架,并为开发基于微生物组的可持续精准作物保护与育种策略提供了重要启示。