导读:
小麦白粉病是由禾本科布氏白粉菌小麦专化型(Blumeria graminis f. sp. tritici, Bgt)引起的真菌病害,是全球小麦生产中最具破坏性的病害之一。培育和推广抗病品种是防控该病害最为经济、有效的策略。经过数十年的系统研究,科研人员已从小麦及其野生近缘种中正式命名了71个抗白粉病基因(Pm基因),其中24个已被成功克隆。这些基因编码的蛋白在结构上高度多样,其作用机制也呈现出复杂性和多样性。
近日,WheatOmics杂志在线发表了四川农业大学、中国农业大学等单位合作撰写的综述“The diversity of cloned wheat powdery mildew resistance genes and the resistance mechanisms”,该综述系统梳理了已克隆Pm基因的来源、染色体定位、遗传进化、蛋白结构、效应因子、抗性机制及其在育种中的应用现状。该综述为深入理解小麦抗白粉病的分子机制、指导抗病育种精准设计提供了重要的理论参考。

研究背景:
传统观点认为,植物抗病基因(R基因)通常编码NLR类蛋白(核苷酸结合富亮氨酸重复结构域蛋白),直接或间接识别病原菌效应子,触发ETI(效应子触发免疫),从而介导植物的抗病性。然而,随着小麦抗白粉病基因研究的不断深入,越来越多的非典型R基因被克隆。这些基因或以成对形式存在的NLR出现(NLR pair),或融合其他功能结构域(NLR-ID),甚至编码与经典NLR结构完全不同的蛋白(如转运蛋白、激酶融合蛋白、膜相关蛋白等)。上述发现不仅拓展了我们对R基因结构多样性的认知,也促使研究者重新审视抗病基因的分类体系及其在育种中的应用策略。
文章内容及主要结果:
1.小麦抗白粉病基因的分类及作用机制
根据所编码蛋白的结构特征及其发挥功能的作用形式,该综述将已克隆的24个Pm基因划分为以下七种主要类型:
单体NLR:单体NLR是抗病基因中最经典的类型,如:Pm1a、Pm2、Pm3b和Pm21等。这类基因通常独立发挥效应子识别功能,可能通过形成类似于拟南芥ZAR1或小麦Sr35的抗病小体钙离子通道,诱导钙离子内流,从而触发细胞死亡和免疫反应,赋予植物抗病性。
成对NLR:两个邻近的NLR基因需协同作用,形成传感器-辅助器模块,分别承担效应子识别和免疫激活功能,介导白粉病抗性。例如Pm5e需与其紧密连锁的RXL共同存在,发挥抗病功能,且二者更倾向形成异源复合物。来自野生二粒小麦的Pm26(TdCNL1/TdCNL5)和Pm68(Pm68-1/Pm68-2)也属于此类。
含整合结构域的NLR(NLR-ID):一些NLR蛋白在进化中融合了其他功能域,这些结构域可能作为“诱饵”参与病原效应子的互作。例如Pm6Sl在CC结构域中整合了锌指BED结构域,形成“CC-BED”嵌合体。Pm52和Pm6则在NLR的C端融合了病程相关激酶结构域(PRK),其LRR和PRK结构域与效应子AvrPm6直接互作。
串联激酶蛋白(TKP):是一类以两个串联激酶结构域为特征的新型免疫受体,近年来在小麦抗病研究中备受关注。例如,从山羊草中克隆的抗白粉病基因WTK4,其N端激酶结构区域含有一个HMA样结构域,能够直接识别白粉菌效应子AvrWTK4。这一发现引发了一个值得深入探讨的问题:TKP中的HMA结构域,是否可视为一种“隐含”的整合结构域(ID),在效应子识别中发挥类似诱饵的功能?
含整合结构域的串联激酶蛋白(TKP-ID):部分抗白粉病TKP(串联激酶蛋白)通过融合其他功能结构域,可能进一步拓展了其效应子识别或信号转导的能力。例如,源自野生二粒小麦的Pm36在C端融合了一个跨膜结构域(TM);源自高大山羊草的Pm57(同时也是Lr9的直系同源基因)则融合了TKP与vWA结构域。然而,这些激酶结构域及整合结构域在抗病相关功能尚不明确,有待进一步研究。
TKP-NLR复合体:Pm24(编码TKP蛋白WTK3)需要与非典型NLR蛋白WTN1协同作用才能激活免疫。值得注意的是,WTK3的等位基因RWT4还能识别稻瘟病菌效应子PWT4,表明TKP-NLR模块可能具有广谱、跨病原识别的潜力。RWT4的N端存在一个部分激酶重复序列(KDup),该结构参与效应子的识别。这引出了两个值得深入探讨的问题:其一,KDup是否可被视为一个整合结构域(ID),在效应子识别中充当诱饵?其二,其他TKP是否也需要依赖NLR协作才能发挥抗病功能?
含整合结构域的激酶(KD-ID):在激酶融合蛋白KFP中,有一类蛋白只包含一个激酶结构域,我们称之为KD-ID。Pm4b编码一个含丝/苏氨酸激酶结构域、多个C2结构域及跨膜结构域的嵌合蛋白,通过可变剪接产生两个转录本,二者对白粉病抗性缺一不可。Pm4b基因也赋予了对麦瘟病的抗性,表明同一个R基因可能识别不同病原菌的不同效应因子。Pm1a的效应因子AvrPm1a/SvrPm4,能抑制Pm4与AvrPm4的互作,该研究揭示了R-Avr互作调控的复杂性。Pm13编码一种混合谱系激酶样(MLKL)蛋白,其N 端含有一个MLKL_NTD结构域,C端含有一个丝氨酸/苏氨酸激酶结构域。然而KFP中的激酶结构域的具体生理生化功能和调控机制不清晰,有待进一步研究。
非典型抗病蛋白:无典型的NLR或激酶类型的抗性蛋白。Pm38(Lr34/Yr18/Sr57)编码ABC转运蛋白,可能通过调节ABA分布或促进细胞壁木质素沉积发挥持久抗性。Pm46(Lr67/Yr46/Sr55)编码己糖转运蛋白,其抗病等位变体丧失糖转运活性,引发离子流变化,改变细胞环境抑制病原菌定殖。感病基因MLO的功能缺失突变也可赋予广谱抗性,通过基因编辑创制的Tamlo-R32材料,实现了抗病与高产的共存,为利用感病基因提供了新范式。
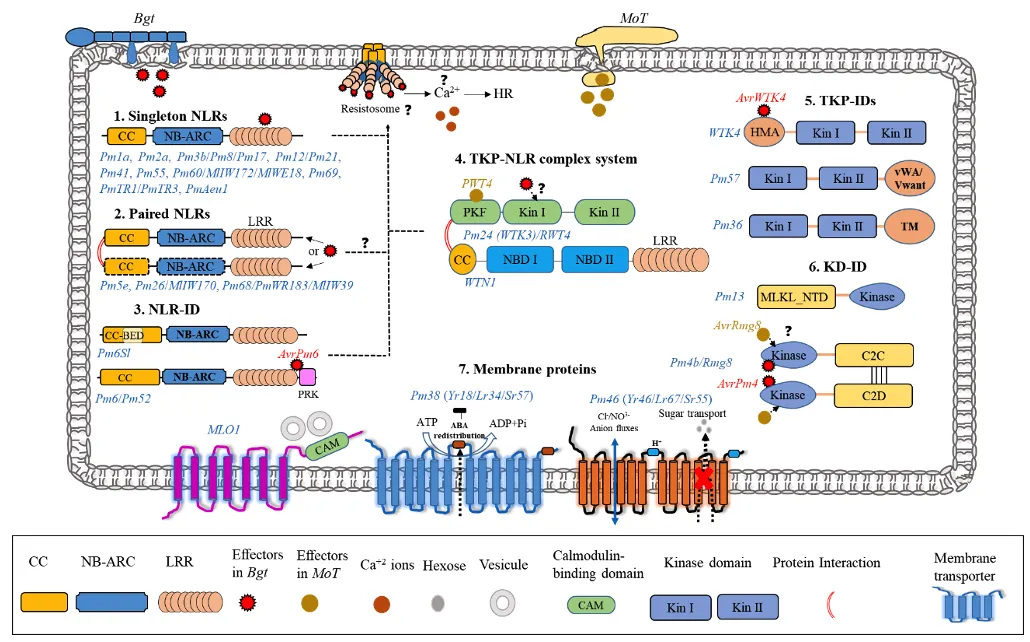 图1:已克隆的小麦抗白粉病基因的结构多样性及其抗病作用机制
图1:已克隆的小麦抗白粉病基因的结构多样性及其抗病作用机制2 小麦抗白粉病基因的生存率及育种现状
基于已发表的毒力频率数据和区域抗性评价,本文梳理了主要抗白粉病基因(Pm基因)的利用现状。随着病原菌的持续进化,Pm3、Pm4a、Pm4b和Pm8等基因的毒力频率较高,表明其抗性已逐步被克服。相比之下,毒力频率较低的Pm基因则可通过与其他抗性基因聚合加以利用,如Pm2+6的组合。此外,部分基因在特定地区已丧失有效性,例如在西澳大利亚,Pm1a、Pm3c、Pm4b和Pm17等已不再具备抗性。而在抗性持久性方面,具有非典型抗性机制的基因表现出明显优势,如转运蛋白型基因Pm38/Lr34,其抗性更难被病原菌突破,具备更持久的田间抗性表现。来源于野生近缘种的部分Pm基因,如Pm12、Pm60、Pm13、PmAeu1和Pm57等,受限于染色体工程育种进展较慢,仍需进一步以提升其育种利用的价值。而另一些基因,如Pm55a,则需要与抑制因子SuPm55共同部署,以维持植株的正常生长发育。总体而言,尽管目前已克隆并鉴定了一定数量的Pm基因,但真正大规模应用于育种的仍十分有限。因此,亟需加强对已知Pm基因,特别是广谱抗性基因的抗性机制研究及育种应用支持,以推动其在作物改良中的有效利用。
3 小麦抗白粉病基因的研究前景与未来展望
整合先进的基因组学与基因编辑工具,对于将源自野生近缘种的R基因高效导入优良作物品种至关重要。借助机器学习、高通量表型分析及前沿测序技术,可快速从种质资源中筛选、鉴定并验证抗性基因。利用图位克隆、全基因组关联分析、突变体筛选、长读长测序技术以及效应因子文库筛选等手段,能够显著加速R基因及其无毒基因(Avr)的克隆进程。深入解析R蛋白与无毒蛋白(R-Avr)的互作机制,将为R基因的工程化改造与设计育种提供新思路。通过基因工程技术改造R基因,使其能够特异性识别病原菌中多个保守效应因子,已成为实现作物广谱抗病性的前沿方向(图2)。此外,结合染色体工程、基因工程及基因聚合育种策略,可有效推动抗病基因在作物改良中的应用。
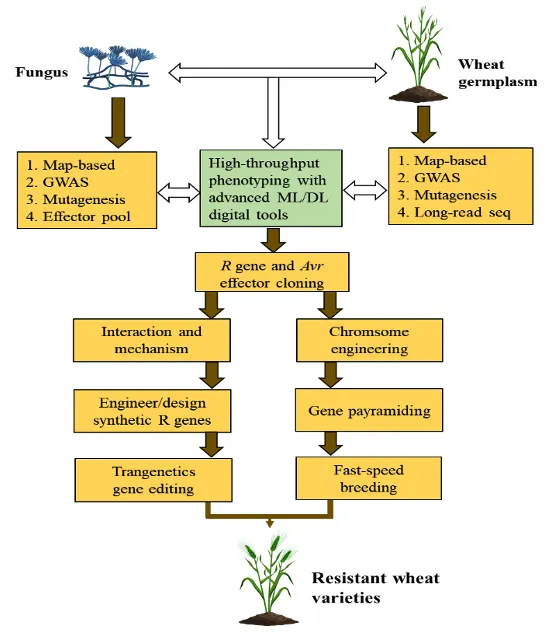 图2:植物抗病基因的发掘与应用的工作流程
图2:植物抗病基因的发掘与应用的工作流程研究团队与项目支持
本研究由四川农业大学、中国农业大学等单位合作完成。四川农业大学小麦研究所李映辉副教授和康厚扬教授为论文的通讯作者。Muhammad Saqlain、陈涛为论文共同第一作者,中国农业大学博士杨祖欢也参与了文章的撰写工作。本论文得到四川省科技计划项目(2024NSFSC1968)、国家自然科学基金项目(32572313)、国家重点研发计划项目(2024YFD1201202)以及四川省科学技术厅项目(2023NSFSC1995、2024NSFSC1968和2025YFHZ0184)的资助。

扫描二维码,免费阅读原文
https://link.springer.com/article/10.1007/s44412-026-00008-8

