2.1 试验材料与接种处理
为探究不同根瘤菌株在相同生境下对宿主共生性能及固氮效率的相对影响,本研究以紫花苜蓿幼苗为试验材料,采用无氮营养液沙培系统进行培养。在幼苗长出第一片真叶的均一发育阶段,分别使用WWL2(登录号MG575945.1)和WE2(登录号MG575932.1)两种苜蓿中华根瘤菌(Sinorhizobium meliloti)悬液对根部进行浇灌接种。试验在严格控制光温湿度的条件下进行,并在接种后21天(成熟根瘤阶段)进行取样。通过测定地上部鲜重以及利用乙炔还原法(ARA)量化固氮酶活性,对两种菌株诱导的植物表型与固氮输出差异进行了生理层面的评估。
2.2 双转录组文库构建与测序分析
为同步解析宿主与共生菌在成熟根瘤微环境中的基因表达动态,研究提取了21天根瘤组织的总RNA,并将其等分为两份以分别构建宿主和共生菌的测序文库。宿主端文库通过富集poly(A)+ mRNA构建,而共生菌端文库则在去除rRNA后构建了保留链特异性的文库。测序在Illumina平台上以双端模式完成。获取原始数据后,通过严格的质量控制滤除低质量序列与接头,随后将宿主和共生菌的纯净reads分别比对至紫花苜蓿参考基因组及Sinorhizobium meliloti 1021模式菌株基因组,从而生成用于后续分析的基因水平计数矩阵。
2.3 差异表达分析与功能验证
为精准识别响应不同菌株接种的分子调控网络,研究基于负二项分布模型对基因计数矩阵进行了TMM归一化处理与差异表达分析,以FDR < 0.05且|log2FC| ≥ 1为阈值筛选差异表达基因(DEGs)。随后,通过GO和KEGG富集分析明确了差异基因的功能分布,并创新性地利用z-score标准化计算了关键功能模块的相对得分,以评估宿主底物交换与共生菌固氮模块间的统计学耦合关系。最后,选取宿主端的ENOD93、SWEET以及共生菌端的nifH、nifD等核心标志基因,以MtACTIN和rpoD为内参基因进行了RT-qPCR定量检测,从而对转录组测序结果的可靠性及提出的分子机制假设进行了严谨的实验验证。
3.1 两种根瘤菌株接种诱导的苜蓿表型差异
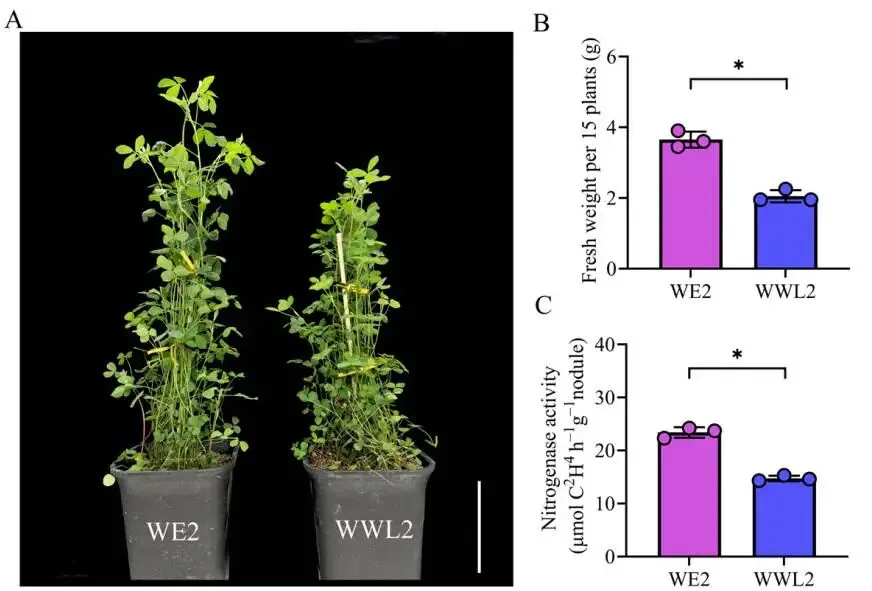
在接种21天后,两种不同的苜蓿中华根瘤菌(WE2和WWL2)诱导了显著的植物表型分化。其中,WE2接种组植株的地上部鲜重和固氮酶活性均显著高于WWL2接种组。基于这种显著的表型差异,进一步对该时期的根瘤组织开展了双转录组测序,以揭示表型背后的分子转录机制(Fig.1)。
图1. 21 dpi根瘤和植株的表型差异(WE2 vs. WWL2)。 (A)接种后21天(dpi)的代表性植物表型;比例尺,10 cm。(B)每个生物学重复的地上部鲜重(g)(每个重复为15株植物的混合样本);n = 3个独立的生物学重复。(C)使用每个生物学重复的混合根瘤通过乙炔还原法(ARA)测定的固氮酶活性;n = 3个独立的生物学重复。数据以平均值 ± SEM表示;圆点表示生物学重复(n = 3)。WE2和WWL2之间的统计学显著性采用双尾非配对Welch's t检验进行评估。表示p* < 0.05。
3.2. 转录组数据的整体分布与样本一致性(PCA)
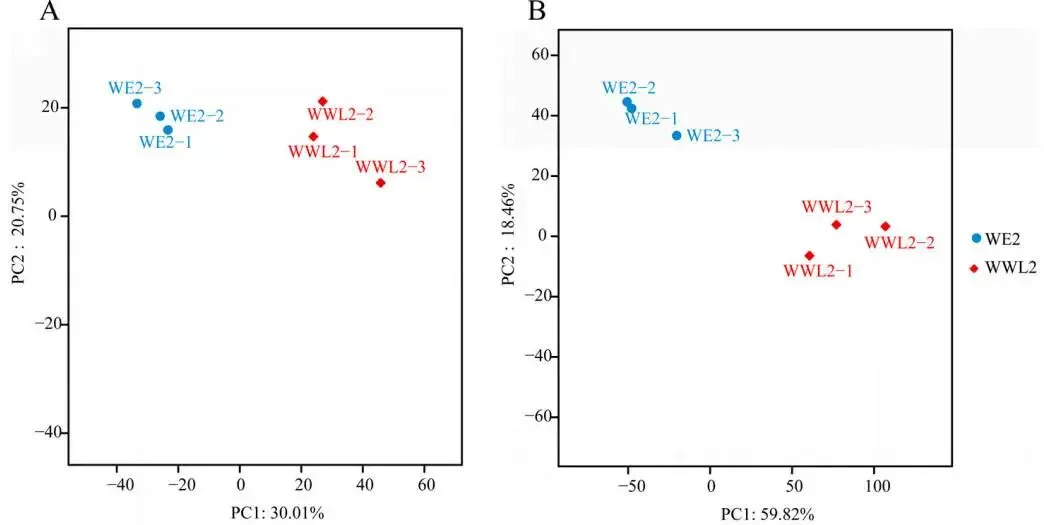
主成分分析(PCA)结果表明,宿主和共生菌的转录组数据均具有极高的生物学重复一致性。同时,WE2和WWL2处理组在主成分空间中呈现出清晰的分离,说明不同菌株的接种在宿主和共生菌双端均诱导了整体转录模式的剧烈重塑,这为后续挖掘差异基因与功能通路奠定了坚实的数据基础(Fig.2)。
图2. 双转录组数据的质量控制。 (A)苜蓿端样本的PCA。(B)根瘤菌端样本的PCA。坐标轴上显示了PC1和PC2解释的方差百分比。
3.3. 宿主(苜蓿)端的差异表达与功能富集(WE2 vs. WWL2)
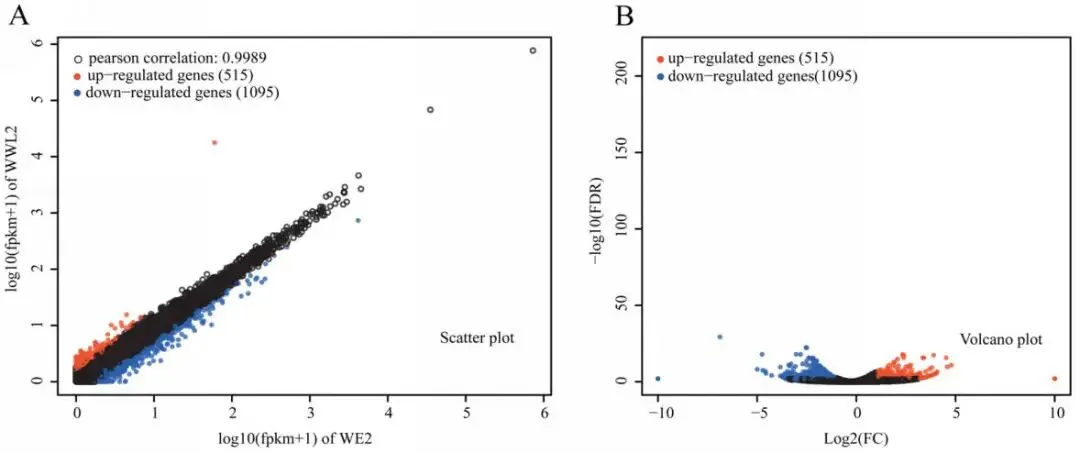
基因上调: 宿主差异基因中,WE2组上调基因数量远多于WWL2组,维持了更广泛的转录程序。
功能富集: 宿主差异基因显著富集于跨膜运输、氧化还原、能量代谢及激素信号传导等通路。
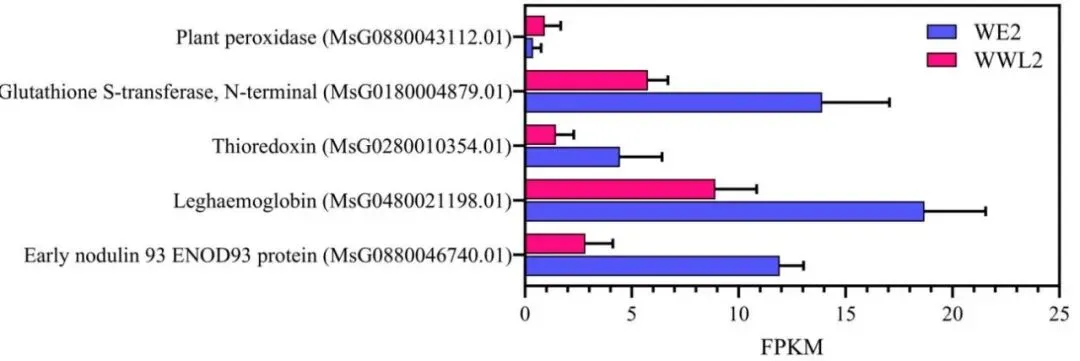
稳态增强:WE2显著上调了成熟根瘤标记基因(ENOD93、leghemoglobin)及SWEET等物质转运与抗氧化基因。
在成熟根瘤阶段,宿主转录组在WE2接种下表现出明显的不对称基因上调趋势。功能富集与聚类分析表明,这些差异表达基因主要集中在物质跨膜运输、氧化还原控制、能量代谢及激素信号传导等关键途径。具体而言,WE2显著增强了宿主端与“底物供应-微需氧稳态”相关的核心模块,包括成熟根瘤标记基因(ENOD93和leghemoglobin)、SWEET家族等糖/氨基酸转运蛋白,以及多种抗氧化与氧化还原调节基因的表达。这一系列协同上调的宿主转录重塑,为解释WE2根瘤中更高效的资源交换与微环境稳态维持提供了坚实的分子证据(Fig.3)(Fig.4)(Fig.5)。
图3. 苜蓿端差异表达概述。 (A)WE2和WWL2的表达水平散点图(log10(FPKM + 1));(B)DEGs的火山图。在这些图中,黑色圆圈代表没有显著差异表达的基因。DEGs的定义标准为FDR < 0.05且|log2FC| ≥ 1;log2FC计算为log2(WWL2 / WE2)。
图4. 根瘤功能和微需氧/氧化还原稳态的宿主模块。 接种苜蓿中华根瘤菌株WE2或WWL2的苜蓿植株在接种后21天收获根瘤(每个处理n = 3个生物学重复)。该图总结了与成熟根瘤功能和微需氧/氧化还原稳态相关的代表性苜蓿DEGs(包括ENOD/leghemoglobin和抗氧化/氧化还原相关基因)。对于每个基因,显示了表达水平(FPKM)和差异表达统计数据(log2FC和FDR);log2FC计算为log2(WWL2 / WE2)(log2FC > 0表示在WWL2中相对于WE2表达更高)。差异表达使用edgeR从原始计数计算得出(见方法)。
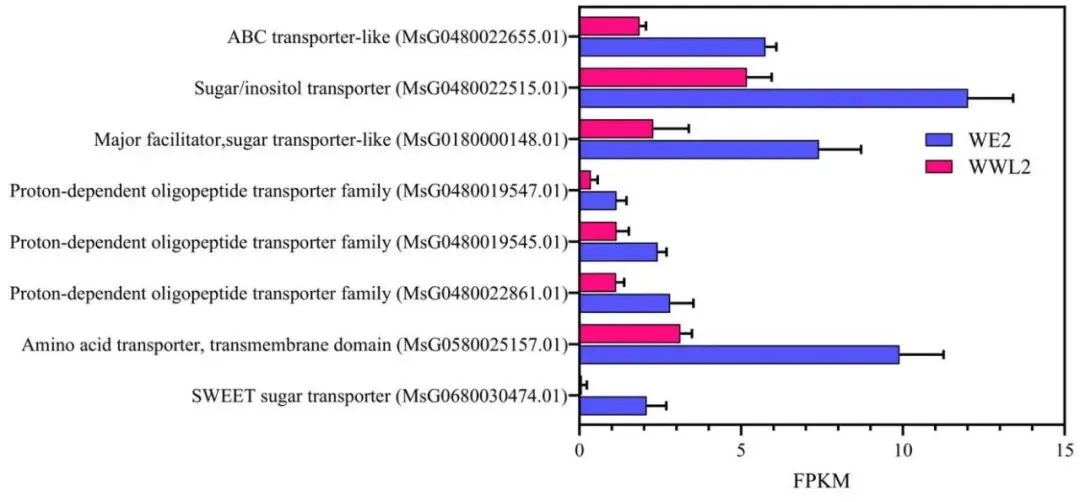
图5. 该图显示了由WE2和WWL2诱导的21 dpi苜蓿根瘤(n = 3)。 显示了与糖转运和氨基酸/肽转运相关的代表性宿主DEGs,并附有FPKM和差异表达指标(log2FC,FDR);log2FC = log2(WWL2 / WE2)。差异表达分析使用edgeR进行。
3.4. 共生菌(根瘤菌)端的差异表达与功能模块(WE2 vs. WWL2)
固氮增强:WE2在固氮(nif)和微需氧呼吸(fix、cbb3)核心模块上的基因表达显著高于WWL2。
适应活跃:WWL2显著上调了环境信息处理、ABC转运系统以及趋化性/运动性(che/fli)相关基因。
策略分歧:WE2偏向于高效的共生固氮输出,而WWL2则停留在营养获取与环境适应的生理状态。
在共生菌端,两种菌株在成熟根瘤内展现出截然不同的转录资源分配策略。WE2显著强化了与共生输出直接相关的核心功能模块,其固氮基因(如nifA、nifH、nifD)和微需氧呼吸及电子传递基因(如fix、cbb3型氧化酶)的表达量远高于WWL2。相反,WWL2则大量上调了与结瘤信号(nod/noe)、表面结构(exo)、趋化性与运动性(che/fli)以及ABC跨膜转运相关的基因。这种转录分歧表明,WE2已完全进入成熟的高效固氮类菌体状态,而WWL2在成熟根瘤期仍将大量能量投入于底物吸收、环境感知与适应性调节中,尚未实现高效的共生功能转化(Fig.6)。
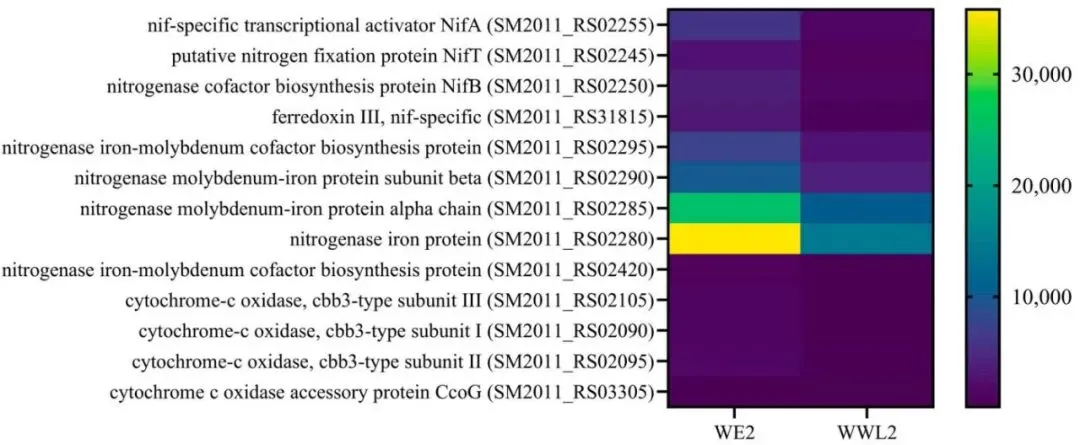
图6. 固氮和微需氧呼吸。 从接种WE2或WWL2的植株中收获21 dpi的根瘤(n = 3)。显示了代表性根瘤菌基因(nif/fdx和cbb3相关的呼吸基因)及其RPKM、log2FC和FDR;log2FC = log2(WWL2 / WE2)。
3.5. 跨物种整合:宿主供应/微需氧稳态与根瘤菌固氮基因表达的协同变化
模块耦合: 宿主的底物供应/微需氧稳态模块与根瘤菌的固氮/微需氧呼吸模块在表达上呈现高度的正相关协同。
实验验证: RT-qPCR验证了ENOD93、SWEET、nifH等核心节点基因的表达趋势,与转录组数据高度一致。
机制模型: 宿主资源供应与微环境调控的增强,完美契合了共生菌固氮程序的激活,解释了表型差异。
通过整合宿主与共生菌双端的关键转录节点,本研究揭示了跨物种的分子协同机制。模块得分与相关性分析表明,宿主的“底物供应/微需氧稳态”模块与根瘤菌的“固氮/微需氧呼吸”模块在WE2接种下发生高度耦合的同步增强(Pearson r = 0.923,p = 0.00868)。RT-qPCR定量实验进一步验证了宿主端ENOD93、leghemoglobin、SWEET以及根瘤菌端nifH、nifD等核心基因表达趋势的可靠性。这种宿主强化氧缓冲与底物输送、共生菌同步激活高效固氮程序的协同模式,完美契合了宏观水平上WE2组更高的固氮酶活性与生物量表型,由此构建了一个解释菌株效应工作框架(Fig.7)(Fig.8)。
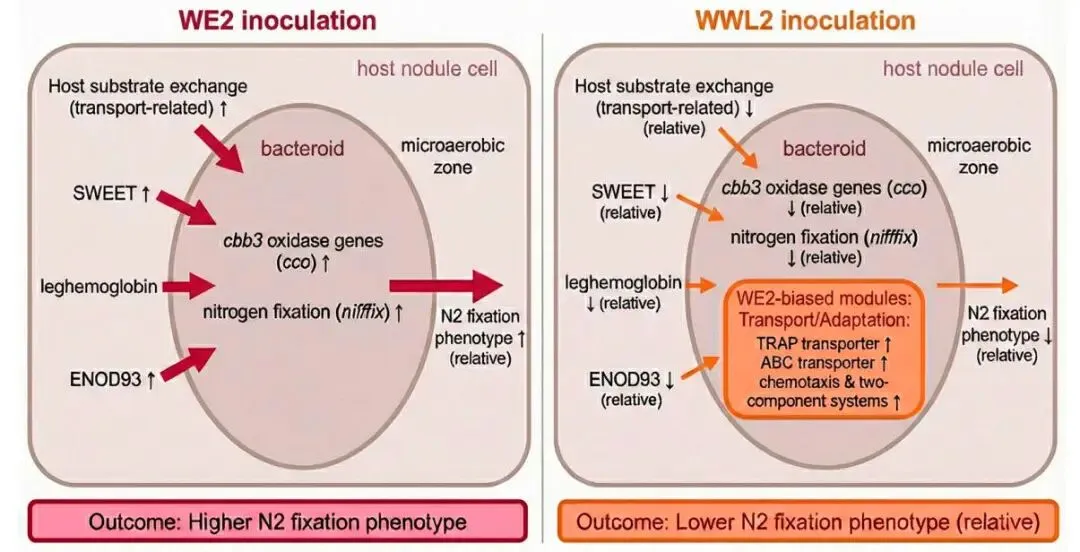
图7. 基于整合双转录组提出的工作框架(示意图)。 该框架突出了在21 dpi成熟根瘤窗口内,与底物交换和微需氧稳态相关的宿主节点(ENOD93、leghemoglobin、SWEET和氨基酸/肽转运)与根瘤菌固氮/微需氧呼吸模块之间的协调变化。该框架用于解释本研究中观察到的表型差异(图1);可以使用固氮酶活性(ARA)等功能指标来测试框架预测(第3.1节;图1C)。↑/↓表示在WE2 vs. WWL2(或反之)比较中相关基因集的上调/下调(21 dpi)。
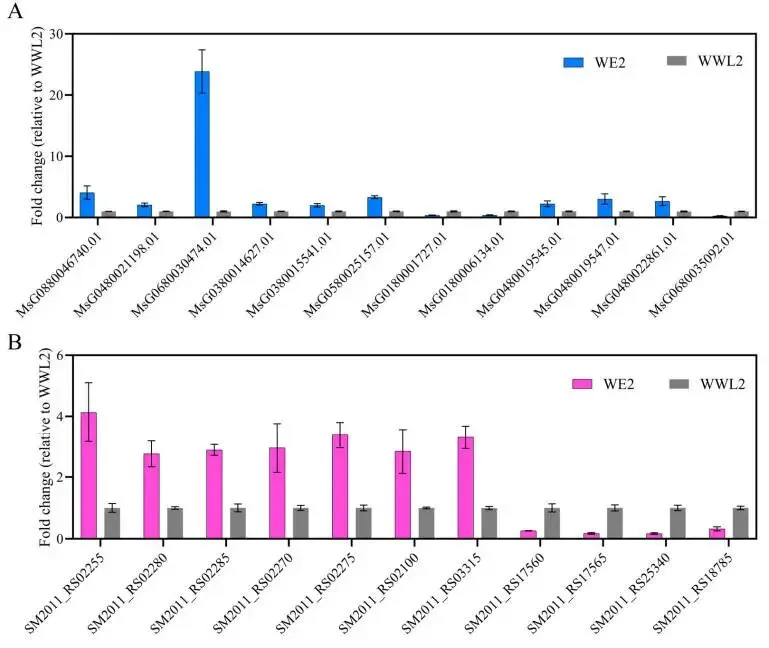
图8. 代表性宿主和根瘤菌基因的RT-qPCR验证。 从接种菌株WE2或WWL2的植株中收获21 dpi的根瘤。(A)宿主基因表达以MtACTIN进行归一化。(B)根瘤菌基因表达以rpoD进行归一化。相对表达量使用2⁻ΔΔCt方法计算。数据以三个生物学重复(n = 3)的平均值 ± SEM表示。每个生物学重复进行三次技术重复测量。







 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
