为提高产环脂肽野生菌贝莱斯芽孢杆菌(Bacillus velezensis)G-1的环脂肽产量和降低其生产成本, 山西农业大学食品科学与工程学院的贾儒健和高振峰以链格孢霉菌为靶标,以廉价菜粕为氮源,以抑菌活性为评价指标,探究了紫外诱变、微波诱变、ARTP(常压室温等离子体)诱变、NTG(亚硝基胍)诱变、DES(硫酸二乙酯)诱变和EMS(甲基磺酸乙酯)诱变6种单一和复合诱变方式对B.velezensis G-1环脂肽合成的影响,旨在建立B.velezensis G-1高产环脂肽突变体的高效诱变方法和筛选出以菜粕为氮源的高产脂肽突变体。结果表明,6种单一诱变方式中化学诱变的正向突变率明显高于物理诱变,且化学诱变中NTG和EMS正向突变率较高,分别为59.97%和59.40%;物理诱变中ARTP正向突变率较高,为31.37%;同单一诱变方式相比,NTG 150 μg/mL、EMS 0.6 mol/L和ARTP 160 s的复合诱变可显著提高B.velezensis G-1的正向突变率,从复合诱变菌株中筛选出1株高产环脂肽突变体,可使环脂肽产量提升2.33倍,离体和活体抑菌活性分别提升52.20%和96.55%。研究结果表明,NTG+EMS+ARTP的复合诱变可成功获取高产环脂肽的B.velezensis G-1突变体,并进一步筛选到了可利用菜粕高产环脂肽且对链格孢霉具有良好抑菌效果的突变体。研究旨在为高产环脂肽B.velezensis G-1突变体筛选及其环脂肽在防治果蔬采后链格孢霉菌侵染的产业化应用奠定理论基础。
链格孢霉菌(Alternaria spp)作为导致水果、蔬菜及冷藏食品腐败变质的主要微生物,环境适应性强,即使在冰温贮藏条件下分生孢子仍具有较强致病性,能产生有毒次级代谢产物链格孢毒素(alternaria toxins),严重威胁食品安全及人体健康[1-2]。因此,在果蔬贮藏期及食品生产过程中防止链格孢霉菌的侵染对果蔬贮藏保鲜和食品安全具有重要意义。
环脂肽主要由芽孢杆菌(Bacillus sp.)经非核糖体途径合成,具有表面活性强、抗植物病原真菌、抗金黄色葡萄球菌、抗癌、抑制黄曲霉毒素合成和降解原油等多种生物学功能[3-4]。研究表明,环脂肽不仅对果蔬采后尖孢镰刀菌(Fusarium oxysporum)、灰葡萄孢菌(Botrytis cinerea)、扩展青霉(Pennicilum expansum)和链格孢菌(Altemuria sp.)等采前、采后病原菌有明显抑制作用,而且活性稳定、靶标位点广、不易产生抗性、安全性高,是一种优秀的生物防腐、保鲜剂开发源[5-10]。然而,环脂肽野生产生菌的产量均较低,且即使通过培养条件优化其产量提高也非常有限,在很大程度上限制了环脂肽的转化应用。
诱变处理是一种在育种界普遍采用的育种方式。目前通过不同诱变处理已快速获得高产脂肽突变菌(UA397、mutHS-407、mutHS-246、mutHS-301、HS-A38)、高产豆豉纤溶酶突变菌株、耐高温产酸菌和富硒纳豆芽孢杆菌等多种突变体,说明诱变育种是提高菌株目标产量简单、有效的途径之一[11-13]。然而,随着菌株诱变研究的不断深入,还发现不同菌株的优选诱变方式及同一菌株不同诱变方式和处理强度间的正向突变率、致死率均存在明显差异,因此,开展不同菌株适宜诱变方式的筛选对获得目的突变体显得尤为重要。
本研究针对目前环脂肽生产中存在的野生菌产量低、发酵成本高和不同菌株诱变育种适宜诱变方式差异较大的问题,以课题组前期鉴选出的对果蔬采后链格孢霉菌具有良好抑制效果的贝莱斯芽孢杆菌(Bacillus velezensis,B.velezensis)G-1环脂肽产生菌为出发菌,采用目前常用的6种物理、化学诱变方式,以环脂肽产量及其抑菌活性为核心评价指标,以廉价菜粕为氮源进行产环脂肽B.velezensis G-1诱变,旨在建立以B.velezensis G-1为出发菌的高产环脂肽突变体优选诱变体系,并筛选出高产环脂肽突变体,为该菌株后期基于廉价菜粕的脂肽物质规模化生产及其在果蔬采后链格孢病害防治中的转化应用奠定基础。
1.1 材料与试剂
B.velezensis G-1和链格孢霉菌,山西农业大学食品科学与工程学院(农产品贮藏保鲜研究所)保藏和提供;胰蛋白胨、酵母浸粉,北京索莱宝科技有限公司;葡萄糖、甲醇、K2HPO4·3H2O、MgSO4·7H2O、MnSO4·7H2O,天津大茂化学试剂厂;亚硝基胍(CAS号:674-81-7),湖北信康医药化工有限公司;硫酸二乙酯(CAS号:64-67-5)、甲基磺酸乙酯(CAS号:62-50-0),上海麦克林生化科技股份有限公司。实验所用试剂均为分析纯。菜粕,四川德阳梦婕菜籽油店。
1.2 仪器与设备
TGL-16M型高速台式冷冻离心机,长沙湘仪离心机仪器有限公司;HZQ-X300型双层摇床,上海一恒科学仪器有限公司;DGL-75G型高压蒸汽灭菌锅,湖南力辰仪器科技有限公司;Easystep-3030型液相色谱仪,上海通微分析技术有限公司;ARTP常压室温等离子体诱变育种仪,洛阳华清天木生物科技有限公司;WSPS-2450-15K型固态微波功率源,成都沃特塞恩电子技术有限公司。
1.3 实验方法
1.3.1 培养基配制
菜粕培养液:菜粕50 g、葡萄糖10 g、K2HPO4·3H2O 0.3 g、MgSO4 ·7H2O 0.2 g、MnSO4·7H2O 0.1 g,蒸馏水1 000 mL,pH值为7.0~7.5,121 ℃湿热灭菌 30 min。
PDA培养基:去皮马铃薯200 g、葡萄糖20 g,蒸馏水1 000 mL,自然pH值,121 ℃湿热灭菌30 min。
LB发酵液:胰蛋白胨10 g、酵母浸粉5 g、NaCl 10 g,蒸馏水1 000 mL,pH值为7.0~7.5,121 ℃湿热灭菌30 min。
1.3.2 菌悬液的制备
1)B.velezensis G-1菌悬液的制备。取1环于-80 ℃甘油保藏的B.velezensis G-1于PDA平板划线,28 ℃活化24 h后,挑取1环接种至LB无菌发酵液中于28 ℃、160 r/min条件下振荡培养24 h获得菌悬液,用于后期诱变实验。
2)链格孢霉菌菌悬液的制备。采用5 mm打孔器打取于PDA平板26 ℃活化7 d后的链格孢霉菌菌饼4个,置于装有5 mL无菌水的10 mL无菌离心管中,涡旋振荡3 min后,4层纱布过滤。通过显微镜血球计数板计数,使用无菌水调整孢子浓度至1.0×105 CFU/mL后备用。
1.3.3 紫外诱变实验
吸取1.3.2节制备的种子菌悬液15 mL加到直径9 cm的无菌培养皿内,使用15 W的紫外灯于垂直距离30 cm处分别照射15、25、35、45、55、65 s进行诱变。将不同诱变处理的菌液稀释106,取0.1 mL稀释液涂布于新鲜的PDA平板上,28 ℃恒温培养24 h,记录菌落数,计算致死率[式(1)]。其次,挑取诱变菌株PDA涂布平板上形态特征与对照存在差异的单菌落于菜粕发酵液中进行发酵培养4 d,并于12 000 r/min离心10 min收集上清液,通过琼脂扩散法以链格孢霉菌为靶标对诱变菌株抑菌活性进行测定,并以突变菌抑菌带高于野生菌3 mm及以上为标准筛选出正向突变体,计算正向突变率[式(2)]。最后,依据正向突变率和致死率筛选出较优紫外诱变时间。每个处理和实验均重复3次。
致死率=(原始平板菌落数-诱变平板菌落数)/原始平板菌落数×100%; (1)
正向突变率=(诱变平板有抑菌活性的菌落数/诱变平板菌落数)×100%。(2)
抑菌活性测定:取20 mL融化后的PDA培养基(硫酸链霉素质量浓度为120 μg/mL)加入90 mm无菌培养中,待其凝固后,取200 μL 1.3.2节中制备的链格孢霉菌菌悬液涂布于表面,待表面干燥后,使用5 mm打孔器打孔,并加入100 μL诱变菌菜粕发酵上清液,置于26 ℃恒温培养5 d后测定抑菌圈大小。
1.3.4 微波诱变实验
微波炉设定最大功率 700 W,脉冲频率2 450 MHz。以30、60、90、120、150、180 s的不同辐照时间对菌悬液进行诱变处理,并在诱变结束后计算致死率和正向突变率,评价不同时间微波诱变处理对B.velezensis G-1的诱变效果。
1.3.5 常压室温等离子体诱变实验
以He作为放电气体,气体流量为10 L/min,射频功率为100 W,等离子体发射源与菌膜之间的距离为2 mm,测定不同诱变时间(20、40、60、80、100、120、140、160、180、200、220、240、260、280、300 s)诱变效果。诱变结束后计算致死率和正向突变率以评价常压室温等离子体(ARTP)不同诱变时间对B.velezensis G-1的诱变效果。
1.3.6 亚硝基胍诱变实验
取1 mL 1.3.2节中制备的 B.velezensis G-1菌悬液与2 mg/mL 亚硝基胍(NTG)母液混合,并使用pH值为6.0的磷酸缓冲液配制成质量浓度分别为50、75、100、150、200、250 μg/mL的系列处理液,于30 ℃水浴30 min,稀释至104终止反应后,计算致死率和正向突变率,评价不同NTG质量浓度对B.velezensis G-1的诱变效果。
1.3.7 硫酸二乙酯诱变实验
取1.3.2节中制备的B.velezensis G-1菌悬液使用体积分数1%的硫酸二乙酯(DES)进行诱变处理。具体的诱变方法:100 mL的三角瓶中加入15 mL pH值为7.0的磷酸缓冲溶液和5 mL菌悬液,再加入0.2 mL DES溶液混匀,30 ℃恒温振荡处理10、15、20、25、30、35、40 min。取1 mL处理液,加入1 mL质量分数25%的Na2S2O3终止反应。然后将处理液适当稀释,取0.1 mL涂布于血平板培养基上,以未经DES处理的菌液稀释涂平板为对照,将处理组和对照组平板于30 ℃培养24~48 h后记录菌落数,并计算致死率。
1.3.8 甲基磺酸乙酯诱变实验
准确称取5.408 g粉末于10 mL容量瓶中,先加入6 mL pH值为7.0的磷酸缓冲液溶解后,定容至10 mL,配成5.0 mol/L母液(4 ℃保藏,临用前用0.45 μm滤膜过滤),随后取1.3.2节中制备的B.velezensis G-1菌悬液与甲基磺酸乙酯(EMS)母液配制成浓度梯度为0.1、0.2、0.4、0.6、0.8 mol/L的混合溶液,混匀处理15 min后,使用0.5 mol/L Na2S2O2溶液终止反应。反应终止60 min后,计算致死率和正向突变率,评价不同EMS浓度对B.velezensis G-1的诱变效果。
1.3.9 复合高效诱变体系建立的响应面优化
通过单因素实验鉴选出的3种诱变方式(NTG、EMS和ARTP)及其较优处理后,为进一步探究3种方式复合诱变是否可提高正向突变率和正向突变效果,采用响应面试验设计了三因素三水平优化试验(见表1)。
表1 响应面试验设计因素与水平
1.3.10 优化复合诱变的B.velezensis G-1高产环脂肽突变体的筛选
以致死率和正向突变率为评价指标,通过响应面试验筛选出较优复合诱变处理后,通过连续继代培养、HPLC、排油圈法、琼脂扩散法分别检测诱变处理后的所有菌株的遗传稳定性、环脂肽产量、排油特性和抑菌活性,筛选出B.velezensis G-1高产环脂肽突变体。
突变体遗传稳定性检测。突变体不经紫外照射连续继代培养5代后,再经紫外照射(最佳诱变时间)后于PDA平板上进行培养和抑菌活性检测,从而确定B.velezensis G-1高产环脂肽突变体的遗传稳定性。
突变体脂肽定性分析。参照高振峰[14]的方法,盐酸沉淀-甲醇抽提的方法提取环脂肽。参照Cherepanova等[3]的HPLC法检测突变体环脂肽峰面积,并通过检测吸收峰组分的排油特性、耐酸碱特性和耐高温特性分析实现吸收峰定性。排油活性测定:取1 mL植物油加入装有15 mL蒸馏水的90 mm培养皿中制备油膜,待油膜稳定后加入10 μL无菌发酵液,观察是否产生排油圈并待排油圈稳定后测定其排油圈大小。酸碱稳定性测定:吸收峰组分首先使用6 mol/L盐酸调节pH值为2,保持30 min,使用4 mol/L NaOH调节pH值为14,保持30 min,最后使用6 mol/L 盐酸将pH值调节为7.0,用于抑菌活性测定。耐高温特性测定:取吸收峰组分置于灭菌锅中经121 ℃湿热处理30 min后,检测其抑菌活性。抑菌活性测定:取20 mL融化后含硫酸链霉素质量浓度为120 μg/mL的PDA培养基加入90 mm无菌培养中,待其凝固后,使用直径为5 mm打孔器,打取1.3.2节中活化后的链格孢霉菌饼,接种至平板中央;然后使用5 mm打孔器打孔,并加入100 μL不同处理环脂肽液体,置于26 ℃恒温培养5 d后观测抑菌带大小。
突变体脂肽定量分析。将1 L发酵液通过盐酸沉淀-甲醇抽提法获得的环脂肽提取物全部经制备液相纯化,并收集活性吸收峰,通过冷冻干燥后称重来实现环脂肽定量。
突变体及其环脂肽抑菌活性测定。1)离体抑菌活性测定方法与1.3.3节相同。2)活体抑菌活性测定参照张晓宇等[15]的方法略加修改。选取大小及成熟度相近的玉露香梨果实,依次经质量分数3% NaClO和体积分数75%乙醇溶液浸泡5 min,无菌水清洗3次,无菌滤纸擦干后,用打孔器在玉露香梨果实表面打直径5 mm、深3 mm的伤口;再取野生菌和突变体以菜粕为氮源的无菌发酵1 000 mL,采用盐酸沉淀-甲醇抽提方法[13]获得其环脂肽提取物,并于冷冻干燥后使用等量(5 mL)无菌水溶解;取野生菌和突变体环脂肽提取液200 μL分3次全部加入玉露香梨果实伤口中,且待伤口处环脂肽液体干燥后使用直径为5 mm的打孔器打取1.3.2节中活化后的链格孢霉菌饼,接种至伤口处,以等量(200 μL)无菌水为对照,每个处理和实验均重复3次;最后,果实置于泡沫果托,使用保鲜膜缠绕封闭保湿后,于25 ℃恒温培养7 d后观测不同处理病斑直径。
1.4 数据处理
采用 SPSS 17.0 Duncan’s 法进行多组样本间差异显著性分析;采用Origin 2018 绘制双Y轴柱-折线状图。
2.1 不同诱变方式诱变效果分析
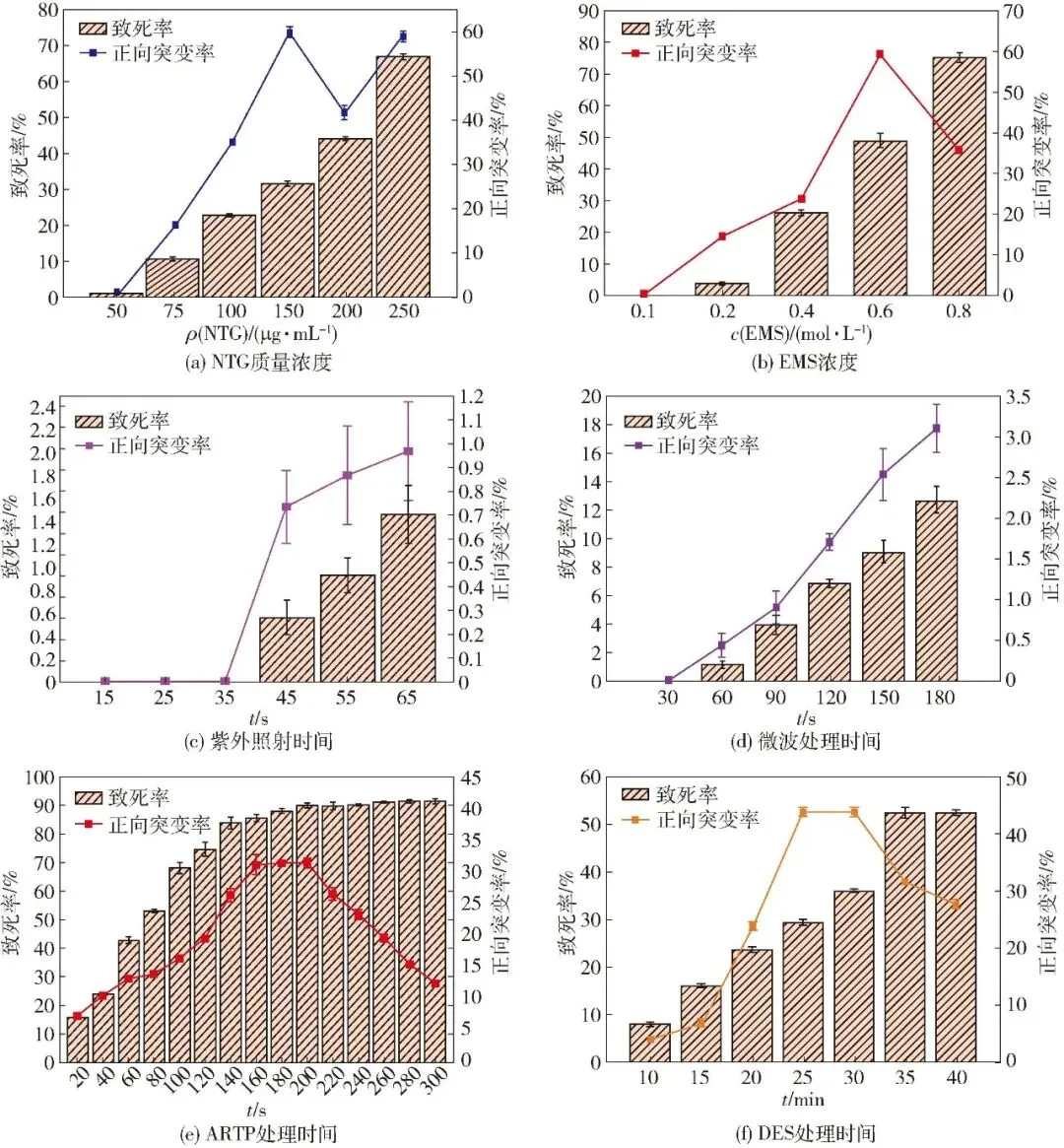
以致死率和正向突变率(突变菌抑菌带同野生菌相比增加3 mm及以上)为评价指标,对不同诱变处理方式的较优突变剂含量或突变时间进行了优化,结果见图1。结果表明:B.velezensis G-1致死率均随着不同处理方式的突变剂含量或突变时间增加呈现逐渐上升趋势,但正向突变率则未表现出相同趋势。随着NTG质量浓度的升高,B.velezensis G-1正向突变率呈现先升高后下降再升高趋势,NTG质量浓度150、250 μg/mL时的正向突变率较高,分别为59.97%和58.97%;但由于250 μg/mL时的致死率较高,因此选择150 μg/mL为较优诱变质量浓度。
图1 不同诱变方式对B.velezensis G-1致死率及正向突变率的影响
随着EMS诱变浓度的增加和ARTP、DES诱变时间的增加B.velezensis G-1正向突变率均呈现先升高后下降趋势,EMS浓度为0.6 mol/L、ARTP诱变时间为160~200 s、DES诱变时间为25~30 min时,B.velezensis G-1正向突变率较高,因此,选择0.6 mol/L、160~200 s和25~30 min为这3种诱变方式较优处理条件。
随着紫外和微波诱变时间的增加,B.velezensis G-1正向突变率呈现逐渐升高趋势,但同其余4种方式相比,正向突变率较低,最高分别仅为0.97%和3.10%,说明这2种物理诱变方式的诱变效果较差,不适用于本实验体系。此外,对6种诱变方式的正向突变效果进行对比后发现,化学诱变中NTG和EMS诱变正向突变率较高,分别为59.97%和59.40%;物理诱变中ARTP诱变正向突变率较高,为31.37%,说明这3种诱变方式在B.velezensis G-1高产菌种创制中具有明显应用优势。
2.2 B.velezensis G-1高效诱变体系建立的响应面优化结果
响应面优化试验结果见表2。结果表明:19种不同组合处理中以处理3和5正向突变率最高,均高于70%,且正向突变率显著高于优化前,但致死率以处理5最低,且同优化前NTG和ARTP单独处理相比具有明显降低;处理1、2正向突变率虽同样高于优化前,但致死率较高。对19种处理正向突变率和致死率结果进行Ⅲ型平方和方差分析,发现二者均模型显著,且失拟项不显著,说明相关模型可用于后续优化诱变条件预测,且结果可靠。
表2 不同诱变组合响应面试验结果
依据响应面试验结果进行二次多项回归拟合,正向突变率=35.06-9.61×A-0.17×B-2.12×C-4.66×AB-2.87×AC-3.89×BC+7.56×A2+6.03×B2+7.70×C2;致死率=75.89+8.46×A+1.25×B+4.37×C-0.56×AB-1.32×AC-1.52×BC-1.24×A2+0.21×B2+0.24×C2。方差分析可以看出3种诱变方式中,NTG对B.velezensis G-1的正向突变率和致死率均存在显著影响,但ARTP仅对致死率存在较大影响。通过比较P值大小可知,3因素对B.velezensis G-1正向突变率的影响从大到小依次为NTG、ARTP、EMS。
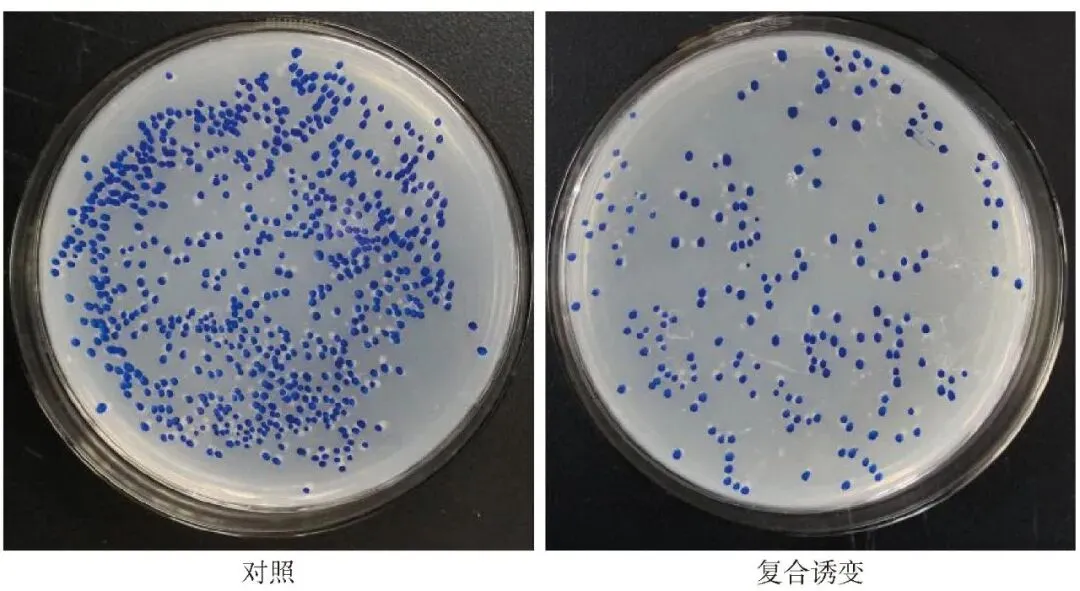
依据建立的正向突变率和致死率回归方程与模型,以致死率最低和正向突变率最高为筛选条件,对B.velezensis G-1较优诱变条件进行优化预测后,选择诱变剂NTG为150 μg/mL、EMS为0.6 mol/L、ARTP诱变时间160 s的复合处理为较优诱变方案,且在该方案下B.velezensis G-1正向突变率和致死率预测值分别为73.58%和64.28%。随后对优选方案进行验证后发现正向突变率和致死率实测值分别为73.63%和64.24%(表2、图2),同预测值相比误差分别为0.07%和0.06%,说明模型预测结果可靠,所得较优方案可较好地提高B.velezensis G-1正向突变率。
图2 化学-物理复合诱变对B.velezensis G-1致死率的影响
2.3 突变体抑菌活性和排油活性分析
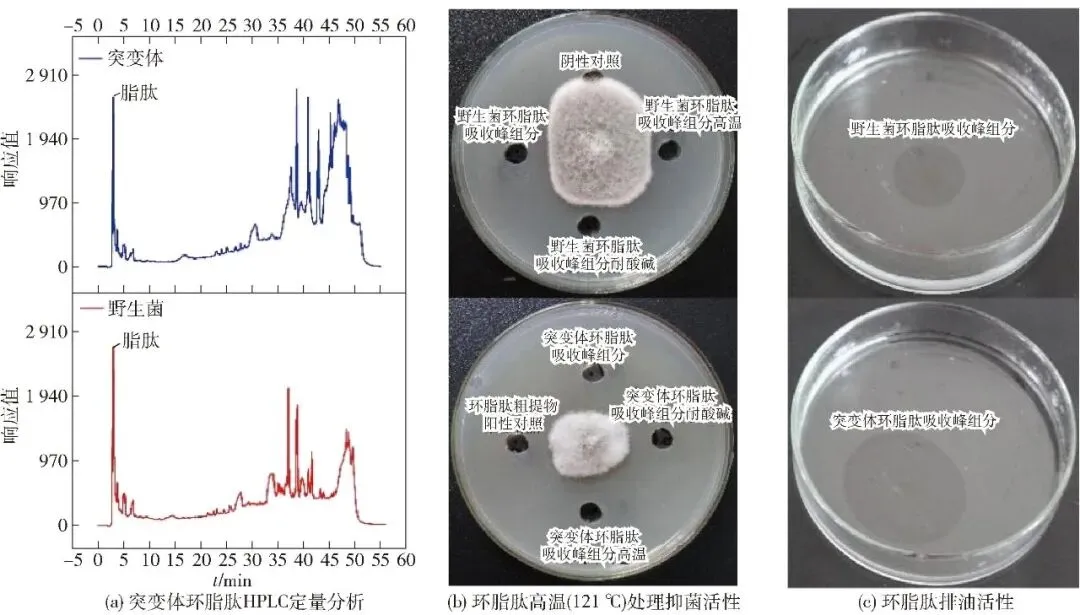
通过发酵实验、平板抑菌实验和排油活性实验相结合的方法从复合诱变平板正向突变体中筛选出1株抑菌活性和排油活性均高于野生菌株的脂肽物质高产突变体(MG16),结果见表3、图3。该突变体的离体抑菌圈和排油圈直径分别达32.45 mm和76.81 mm,同对照组(抑菌圈21.32 mm、排油圈42.17 mm)相比,抑菌活性和排油活性分别提升了52.2%和82.1%。
图3 突变体环脂肽含量及环脂肽抑菌活性、耐高温特性、耐酸碱特性和排油活性分析
表3 突变体菌株和野生菌株的抑菌活性、排油活性及环脂肽产量
同列不同小写字母表示组间数据差异显著(P<0.05)。
通过环脂肽物理特性分析明确突变体环脂肽吸收峰HPLC出峰位置后,对突变体和野生菌基于菜粕氮源的环脂肽产量进行了定量分析。结果表明:突变体环脂肽产量比野生菌提升了2.33倍;相同条件下突变体环脂肽吸收峰的抑菌带、排油圈和经高温、酸碱处理后的抑菌活性均高于野生菌,充分说明突变体可稳定高产环脂肽。
此外,对突变体菌株及其环脂肽的活体抑菌活性进行测定,结果见图4。结果表明:同野生菌相比,突变体菌株和同等制备条件下的环脂肽的活体抑菌活性均显著优于野生菌,且同野生菌相比抑菌活性分别提升了25.21%和96.55%,再次证明复合诱变显著提高了突变体的环脂肽产量。
不同小写字母表示病斑直径差异显著(P<0.05)。
图4 突变体菌株活菌及其环脂肽抑菌活性
随着环脂肽生物作用研究的不断深入,其在食品和医药领域的防腐、抗菌功效已逐渐被人们广泛认可,但大量研究表明野生菌株的环脂肽产量均较低,难以满足转化应用后的市场需求,因此,如何提高环脂肽产量成为脂肽物质走向市场化应用前急需突破的关键问题。另外,尽管近年来分子育种技术取得了巨大进展,但迄今为止诱变育种仍是微生物育种和种业育种中的核心技术手段,且利用该技术已成功选育出了一些高产目标物质的微生物突变体(如富硒纳豆芽孢杆菌[13]、低产杂醇高产酯酵母菌株[16]、高产脂肽芽孢杆菌[17-20]),并发现不同菌株和不同属种微生物的高效正向诱变方式存在一定差异,因此,鉴选不同菌株适宜的诱变方式对高效诱变育种具有重要意义。
针对B.velezensis G-1环脂肽产量低、成本高的问题,本研究以菜粕培养液为营养源,以诱变育种为核心技术,以课题组鉴选出的产环脂肽且对果蔬采后链格孢霉菌具有良好抑菌效果的B. velezensis G-1为出发菌,选择了目前常用的3种物理和3种化学诱变方式,以链格孢霉菌为靶标菌,以正向突变率(突变体抑菌带高于野生菌3 mm及以上)为评价指标,通过单因素实验对比分析了不同诱变方式的诱变效果。结果发现,不同诱变方式中化学诱变正向突变普遍较高,但致死率亦同样高于物理诱变。3种物理诱变方式中ARTP的诱变效果较好,优选诱变时间为160~200 s,平均正向突变率为31.37%,显著高于紫外和微波诱变;3种化学诱变方式中NTG和EMS诱变处理正向突变率较高,二者在优选诱变含量为150 μg/mL和0.6 mol/L时的正向突变率分别为59.97%和59.40%。辛磊等[18]紫外诱变Bacillus subtilis XL05,突变体的抑菌活性显著高于野生菌;谢定刚等[19]紫外诱变枯草芽孢杆菌mutHS-301,突变体的抑菌活性和脂肽产量显著高于野生菌;陆雅琴[20]紫外诱变芽孢杆菌P1和P6,突变体的脂肽产量显著高于野生菌;吴红玉[21]紫外诱变解淀粉芽孢杆菌Ht-q640s时的正向突变率较高。这些研究结果与本研究存在明显差异,说明不同菌株的较优物理诱变方式及处理时间存在一定差异。李光等[22]研究发现,Bacillus subtilis E7 ARTP诱变突变体的surfactin产量显著高于野生菌;吴越等[23] 诱变Bacillus subtilis SF90-5 ARTP,突变体抑菌活性和表面活性素产量显著高于野生菌;靖雅丽[24]研究表明,Bacillus sp.N110 ARTP诱变突变体的表面活性素产量显著高于野生菌,这些研究结果与本研究相似,证明ARTP诱变可提高芽孢杆菌脂肽物质产量和抑菌活性,但相关研究均未进行相关菌株的较优诱变方式筛选,不能说明ARTP为相关菌株的优选诱变方式。本研究化学诱变研究结果同孟攀攀[25]确定的枯草芽孢杆菌fmbJ较优诱变剂含量分别为800 μg/mL和1.0 mol/L,张莉[26]确定的枯草芽孢杆菌BS1 NTG的优选诱变剂质量浓度为0.4 mg/mL,闫冬[27]确定的多粘类芽孢杆菌JSa-9的NITG较优诱变质量浓度为50 μg/mL存在一定差异,这些差异的产生与菌株个体差异关系密切,说明不同菌株的优选诱变方式、诱变剂含量和诱变时间存在较大差异。
微生物诱变选育过程中,单一诱变往往会产生疲劳效应,采用多种诱变选育相结合的复合诱变通常能够获得更加高产的菌株,因此,在明确 B. velezensis G-1的较优诱变方式及较优处理浓度或时间后,为进一步探究不同诱变方式的复合诱变效果,本研究采用响应面试验对单因素优选诱变方式的较优复合诱变处理进行了系统优化。结果表明,B. velezensis G-1的优选复合诱变处理为NTG 150 μg/mL、EMS 0.6 mol/L和ARTP 160 s,且从该诱变方式下筛选出1株突变体,其同野生菌相比以菜粕为氮源时的环脂肽产量提升了2.33倍,离体和活体抑菌活性分别提升了52.20%和96.55%。相关研究结果同张莉[26]、高姚建等[28]的复合诱变方式存在明显差异,再次证明不同菌株适宜的诱变方式不同。另外,本研究获得的突变体环脂肽产量提升效果同孟攀攀[25]、张莉[26]的提升效果存在一定差异,这种差异的产生可能与出发菌株自身因素及培养液配方、发酵条件存在较大差异有关,且本研究并未针对突变体进行发酵条件优化,后期还需在相同发酵条件下同已报道菌株对比环脂肽产量提升效果,以确定B. velezensis G-1同其他菌株相比是否存在明显应用优势。另外,本研究还发现B. velezensis G-1菌株及其环脂肽的离体和活体抑菌效果存在明显差异,而目前在果蔬采后拮抗微生物筛选上还以离体筛选为主,因此从生产应用实际出发,建议增加活体筛选甚至贮藏实验来避免假阳性菌株的出现。
本研究以廉价菜粕为氮源,通过单因素和响应面优化相结合的方式,证明了复合诱变效果优于单一诱变,鉴选出了适宜B. velezensis G-1的NTG (150 μg/mL)+EMS (0.6 mol/L)+ARTP (160 s)三级复合诱变方式及可以菜粕为氮源的高产环脂肽突变体;获得的突变体同野生菌相比,环脂肽产量提升了2.33倍,离体和活体抑菌活性分别提升了52.20%和96.55%。然而,其他诱变方式对B. velezensis G-1的诱变效果(LiCl、抗生素、亚硝酸等)及其突变体环脂肽对其他果蔬采后致腐病原的抑菌效果、高产环脂肽分子机制与已报道的环脂肽产生菌的差异等相关研究还需进一步探究。研究旨在筛选出1株生产成本较低的高产环脂肽突变体,为其在果蔬采后病害防治的产业化应用奠定基础。
引用格式:贾儒健,高振峰.产环脂肽Bacilus velezensis G-1高效诱变育种及对链格孢霉的抑菌效果[J]. 食品科学技术学报,2025,43(2):95-105.JIA Rujian,GAO Zhenfeng. Efficient breeding of cyclic lipopeptide producing Bacillus velezensis G-1 by mutagenesis and its antifungal activity against Alternaria alternata[J]. Journal of Food Science and Technology, 2025,43(2):95-105.基金项目:山西省应用基础研究计划项目(20210302124512)。Foundation:Fundamental Research Program of Shanxi Province (20210302124512).
制作:张羽
编辑:张逸群、李宁
审核:叶红波











 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
