合作文章丨安徽农业大学团队揭示了TaDof-2D-miR1832-TaP450-7A调控模块介导低温诱导小麦种子休眠解除的核心机制
- 2026-08-03 05:51:51

英文题目:The TaDof-2D-miR1832-TaP450-7A module regulates low temperature-induced release of seed dormancy in wheat
中文题目:TTaDof-2D-miR1832-TaP450-7A模块调控小麦低温诱导种子休眠解除
期刊名称:Plant Communications
影响因子:11.6
作者单位:安徽农业大学农学院
普奈斯提供服务:赤霉素组分检测
DOI号:https://doi.org/10.1016/j.xplc.2026.101749
研究内容
该研究以强休眠小麦地方品种“歪头白”及多个不同休眠水平的小麦品种为试验材料,整合全转录组测序、分子生物学实验、生理生化检测及遗传验证等,系统探究了TaDof-2D-miR1832-TaP450-7A调控模块介导低温诱导小麦种子休眠解除的分子机制与应用价值。研究聚焦小麦种子发育中后期(开花后21~35天)的低温响应过程,通过设置低温(15℃日/10℃夜)与常温(25℃日/20℃夜)处理,结合转基因技术(过表达、沉默株系构建)、突变体分析(甲基磺酸乙酯诱变)、分子互作验证(酵母单杂交、电泳迁移率变动分析、双荧光素酶报告基因实验)等方法,系统解析了调控网络中关键基因的功能及互作关系。
研究发现,低温处理可显著下调新型miRNA(miR1832)的表达,该基因正调控小麦种子休眠,其过表达能增强种子休眠,沉默则抑制休眠;miR1832启动子区-670bp处的A/G单核苷酸突变与不同小麦品种的休眠表型显著关联,A等位基因与强休眠性、抗穗发芽特性紧密相关,且该等位基因在小麦地方品种中分布频率较高(76.6%),但在改良品种中极少存在(1.6%)。低温响应型Dof转录因子TaDof-2D可特异性结合miR1832启动子的A等位基因位点,抑制其转录表达;而miR1832通过靶向切割细胞色素P450基因TaP450-7A(负调控种子休眠)抑制其表达,TaDof-2D通过抑制miR1832的表达,解除其对TaP450-7A的靶向抑制,进而调控种子休眠。
生理生化分析表明,该调控模块通过影响α-淀粉酶活性及脱落酸、赤霉素的生物合成与信号通路介导低温诱导的种子休眠解除:低温处理下,TaDof-2D表达上调抑制miR1832,促进TaP450-7A表达,进而上调赤霉素生物合成相关基因(TaGA20ox1、TaGID1-1B/1D等)和脱落酸分解代谢基因(TaABA8'OH2-5B/5D等)的表达,下调脱落酸信号通路基因(TaABI5-3B/3D等)和α-淀粉酶抑制基因(TaCM1-7D、TaCM2-7B)的表达,最终导致种子中赤霉素(GA1、GA3)含量升高、脱落酸含量降低、α-淀粉酶活性增强,推动休眠解除。此外,基于miR1832启动子A/G突变开发的CAPS分子标记1832,可有效区分不同休眠水平的小麦品种,为抗穗发芽小麦品种的分子育种提供了实用工具。
该研究明确了TaDof-2D-miR1832-TaP450-7A模块在温度响应型种子休眠调控中的核心作用,揭示了小麦应对低温胁迫的分子适应机制,同时为通过分子设计育种培育气候适应性增强的小麦品种提供了关键基因靶点与技术支撑。
研究路线
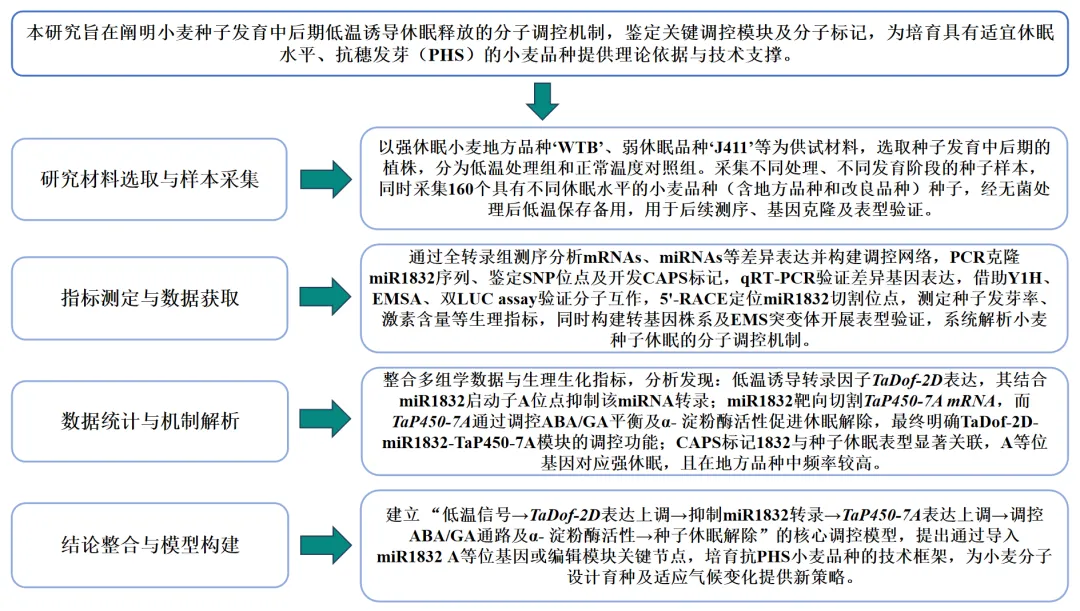
样本处理
强休眠小麦地方品种“歪头白”于2017-2018生长季在合肥(31°58′N,117°24′E)自然田间条件下种植,试验田全程无病虫草害。小麦开花前进行常温(NT)或低温(LT)处理,随后进行全转录组测序:选取60个代表性主茎穗标记开花时间,于开花后21~35天将植株转移至人工气候箱(16h光照/8h黑暗,相对湿度70%),分别进行低温(10℃夜/15℃日)或常温(20℃夜/25℃日)处理,于开花后21天(21DPA-NT)和35天(35DPA-NT、35DPA-LT)收获种子,液氮速冻后于-80℃保存,用于RNA提取。
选取4个小麦品种[强休眠品种“歪头白”(WTB)、“红芒春21”(HMC21);弱休眠品种“中优9507”(ZY9507)、“京411”(J411)]用于miR1832的克隆和表达分析;于2021-2022生长季在合肥种植124个小麦地方品种和245个改良品种;选取160个不同休眠水平的小麦品种,用于验证miR1832启动子区-670bp处A/G突变与种子休眠表型的关联性,上述160个品种于2021-2022和2022-2023生长季在合肥、淮北、宿州、郭河种植,且主要来源于我国黄淮麦区、长江中下游麦区、北方麦区和西南麦区。
小麦野生型“菲尔徳”(Fielder)、miR1832过表达株系、miR1832沉默株系、“Fielder”背景的TaDof-2D过表达株系(1832-G基因型)和TaP450-7A过表达株系,于2023-2024生长季在石家庄河北省农林科学院堤上试验站(37°56′N,114°43′E)种植,选取纯合T2代转基因植株用于试验;miR1832与TaP450-7A过表达杂交株系(pre-miR1832-OE#4/P450-7A-OE#7)及“WTB”背景的TaDof-2D过表达株系(Dof-2D-A-OE,1832-A基因型)在人工温室中种植(23±2℃,8h黑暗/16h光照);利用“J411”背景的甲基磺酸乙酯诱变突变体(dof-2d、p450-7a)验证TaDof-2D和TaP450-7A对种子休眠的调控作用,为减少遗传背景突变的干扰,将突变体与“J411”回交两代后自交,获得BC2F3代后代用于试验。
拟南芥野生型“哥伦比亚-0”(Col-0)及转基因T2代植株经4℃低温处理3天后,转移至人工温室(24±1℃,16h光照/8h黑暗)培养7天,随后移栽至蛭石与黑土体积比3:1的方形花盆中;水稻野生型“日本晴”(Nip)及转基因T2代植株在人工温室中种植(28℃日/25℃夜,10h光照/14h黑暗,相对湿度70%,光照强度200μmol光子·m-2·s-1)。
研究结果
01
种子发育中后期的低温诱导种子休眠解除
为探究种子发育中后期低温对小麦种子休眠的影响,本研究以强休眠小麦地方品种“WTB”为材料,在其开花后21~35天进行低温(15℃日/10℃夜)或常温(25℃日/20℃夜)处理,于开花后35天收获种子并测定含水量,结果显示常温处理种子含水量为12%,低温处理种子为23%。萌发实验表明,低温处理种子的发芽率达100%,而常温处理种子发芽率为0%(图1A-C)。为明确种子休眠表型的变化由低温本身还是含水量差异引起,将低温处理种子自然风干至含水量约12%,萌发实验显示其发芽率仍达100%,说明低温是诱导种子休眠解除的主导因素,而非含水量增加。此外,低温处理还显著改变了种子中脱落酸、赤霉素的含量及α-淀粉酶活性:与常温处理相比,低温处理种子中赤霉素GA1和GA3含量显著升高,脱落酸含量显著降低,α-淀粉酶活性显著增强(图1D-F)。以上结果证实,小麦种子发育中后期遭遇低温可诱导其休眠解除。
02
miR1832作为调控网络核心因子参与低温诱导的种子休眠解除
为解析低温诱导小麦种子休眠解除的分子机制,本研究对常温和低温处理的“WTB”脱壳种子进行全转录组测序,共鉴定出4862个差异表达基因(DEGs),其中低温处理组相较于常温处理组有4010个基因上调表达、852个基因下调表达。KEGG通路富集分析显示,这些DEGs显著富集于碳水化合物代谢、能量代谢、氨基酸代谢、信号转导等生物学过程。同时,还鉴定出82个差异表达miRNAs、108个差异表达长链非编码RNA(lncRNAs)和9个差异表达环状RNA(circRNAs)。通过实时荧光定量PCR验证了随机选取的40个mRNAs、10个miRNAs、20个lncRNAs和5个circRNAs的表达模式,结果与转录组测序一致,证实了测序结果的可靠性。
为明确参与低温诱导种子休眠解除的miRNAs与靶标基因的功能关系,本研究基于差异表达的mRNAs和miRNAs构建调控网络,预测结果显示二者存在一对一、一对多和多对一的调控关系。在该调控网络中,miR1832(unconservative_chr1A_1832)处于核心节点且受低温下调表达(图1G);实时荧光定量PCR(qRT-PCR)验证了miR1832在常温与低温条件下的表达模式,证实其在低温诱导的种子休眠解除过程中发挥核心作用。
03
miR1832在常温条件下也参与种子休眠的调控
拟南芥DOG1、小麦TaMFT-3A、水稻SD6等休眠相关基因在常温和低温条件下均参与种子休眠调控。为明确miR1832在常温下是否调控种子休眠,本研究检测了其在两个强休眠品种(“WTB”、“HMC21”)和两个弱休眠品种(“ZY9507”、“J411”)种子发育阶段(开花后21、28、35天,休眠建立阶段)、后熟阶段(收获后7、14、21天,休眠解除阶段)和吸胀阶段(吸胀1、6、9、12、36小时,休眠解除阶段)的表达模式。结果显示,miR1832的表达量在种子休眠建立过程中升高,在休眠解除过程中降低;且在整个过程中,强休眠品种“WTB”和“HMC21”中miR1832的表达量始终显著高于弱休眠品种“ZY9507”和“J411”。以上结果表明,miR1832在常温条件下参与小麦种子休眠的建立与维持。
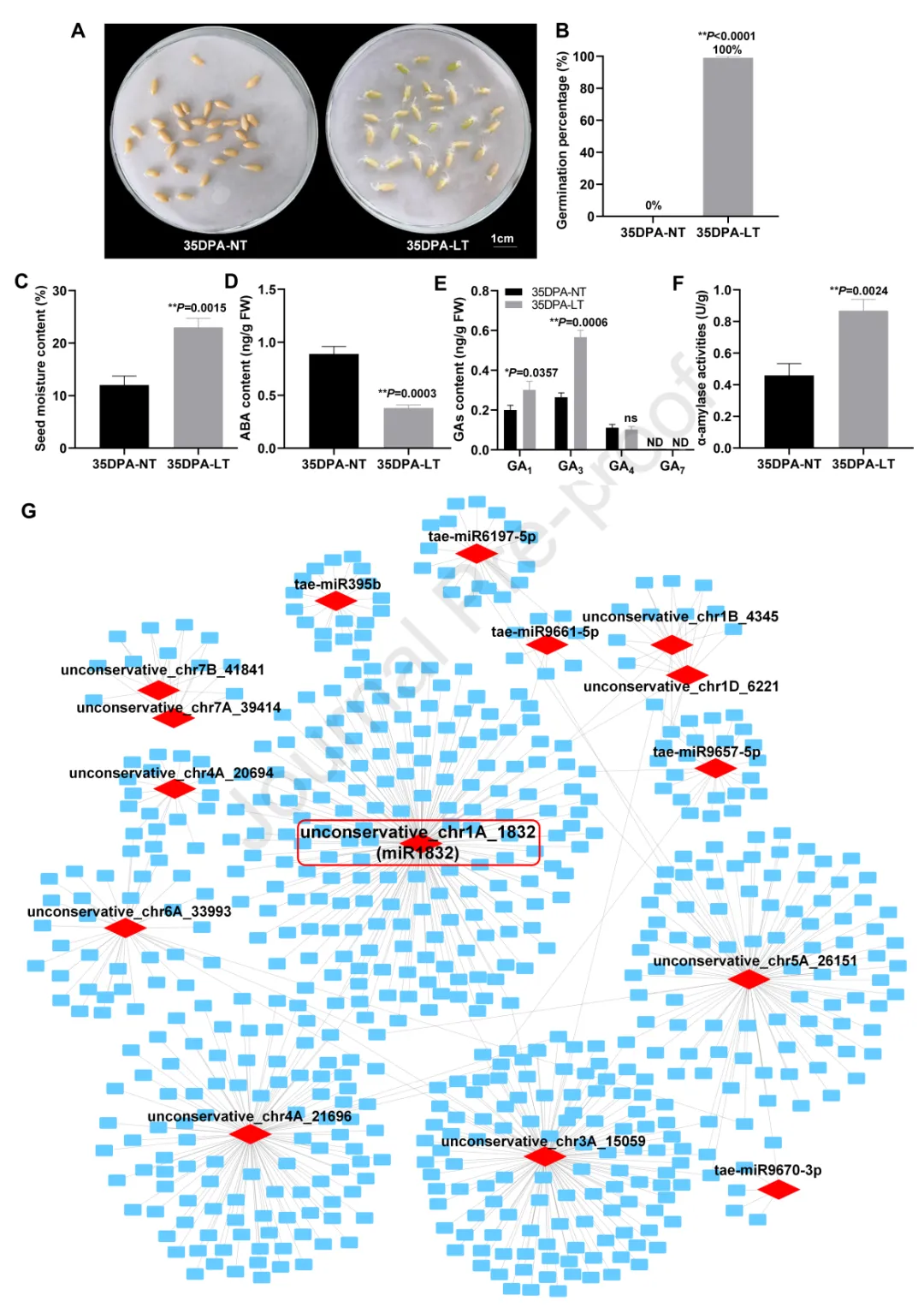
为验证miR1832对小麦种子休眠的调控作用,本研究在小麦野生型“Fielder”中过表达miR1832的前体序列,并沉默其成熟转录本,分别获得3个过表达株系(pre-miR1832-OE#3、pre-miR1832-OE#4、pre-miR1832-OE#7)和3个短串联靶标模拟物沉默株系(miR1832-STTM#2、miR1832-STTM#3、miR1832-STTM#4)。表达检测显示,与“Fielder”相比,过表达株系中miR1832的表达量显著上调,而沉默株系中其表达量显著下调。萌发实验表明,3个过表达株系的发芽率(pre-miR1832-OE#3为47%、pre-miR1832-OE#4为35%、pre-miR1832-OE#7为42%)显著低于“Fielder”(84%),而3个沉默株系的发芽率(miR1832-STTM#2为97%、miR1832-STTM#3为95%、miR1832-STTM#4为96%)显著高于“Fielder”(图2A-C)。以上结果证实,miR1832正调控小麦种子的休眠性。
植物中同源的休眠相关基因通常具有保守的功能,如拟南芥AtABI3与小麦TaVp-1、拟南芥DOG1与小麦TaDOG1L1、水稻OsSdr4与小麦TaSdr、大麦HvMKK3与小麦TaMKK3-A、大麦HvQsd1与小麦TaQsd1、水稻OsSD6与小麦TaSD6等。序列比对显示,小麦miR1832与水稻同源基因OsmiR1832的相似性达89%,与拟南芥同源基因AtmiR1832的相似性达78%。为探究miR1832、OsmiR1832和AtmiR1832在种子休眠调控中的功能保守性,本研究在水稻和拟南芥中异源过表达小麦miR1832的前体序列,并通过短串联靶标模拟物沉默OsmiR1832和AtmiR1832的成熟转录本。萌发实验表明,过表达miR1832的水稻和拟南芥种子发芽率显著低于野生型“Nip”和“Col-0”,而沉默OsmiR1832和AtmiR1832的种子发芽率显著高于野生型。以上结果表明,miR1832可跨物种增强植物种子的休眠性,且水稻OsmiR1832和拟南芥AtmiR1832与小麦miR1832具有相似的功能。
04
miR1832启动子区的A/G突变与种子休眠表型差异显著相关
为探究miR1832介导小麦种子休眠表型差异的分子基础,本研究从强休眠品种“WTB”、“HMC21”和弱休眠品种“ZY9507”、“J411”中克隆了miR1832上游1000bp的启动子序列及前体序列。结构分析显示,miR1832具有典型的茎环结构;序列比对发现,强休眠品种与弱休眠品种的miR1832启动子和前体区存在两个单核苷酸多态性突变。启动子顺式作用元件预测分析表明,miR1832启动子区-670bp处的A→G突变,可能导致弱休眠品种“J411”和“ZY9507”中该位点的Dof蛋白结合基序缺失(图2D);而前体区+156bp处的T→C突变未改变其茎环结构。
为验证miR1832启动子区-670bp处的A/G突变是否影响种子休眠表型,本研究基于该突变开发了酶切扩增多态性序列标记(CAPS-1832)(图2E),并利用该标记对160个不同休眠水平的小麦品种进行基因分型,将该标记的两个等位基因命名为1832-A和1832-G。U检验结果显示,1832-A和1832-G基因型小麦品种的发芽指数存在极显著差异(图2F),其中1832-A等位基因与低发芽指数(强休眠、抗穗发芽)显著关联,1832-G等位基因与高发芽指数(弱休眠、易穗发芽)显著关联(P<0.01)。为进一步验证该A/G突变与种子休眠表型的关联性,本研究检测了10个1832-A基因型和10个1832-G基因型小麦品种中miR1832的表达量,结果显示1832-A基因型品种中miR1832的表达量显著高于1832-G基因型品种。综上,miR1832启动子区的A/G突变与小麦种子休眠表型差异显著相关,其中A等位基因与强休眠性和抗穗发芽性关联。
为探究与强休眠、抗穗发芽相关的1832-A优异性等位基因在小麦育种中的利用情况,本研究利用CAPS-1832标记对124个小麦地方品种和245个改良品种进行基因分型。结果显示,在124个地方品种中,1832-A等位基因的分布频率达76.6%(95个),显著高于1832-G等位基因(23.4%,29个);而在245个改良品种中,仅4个品种(1.6%)携带1832-A等位基因,其余241个品种(98.4%)均为1832-G等位基因(图2G)。以上结果表明,1832-A等位基因在现代小麦育种中未得到充分利用,可作为分子设计育种中改良小麦穗发芽抗性的候选等位基因。
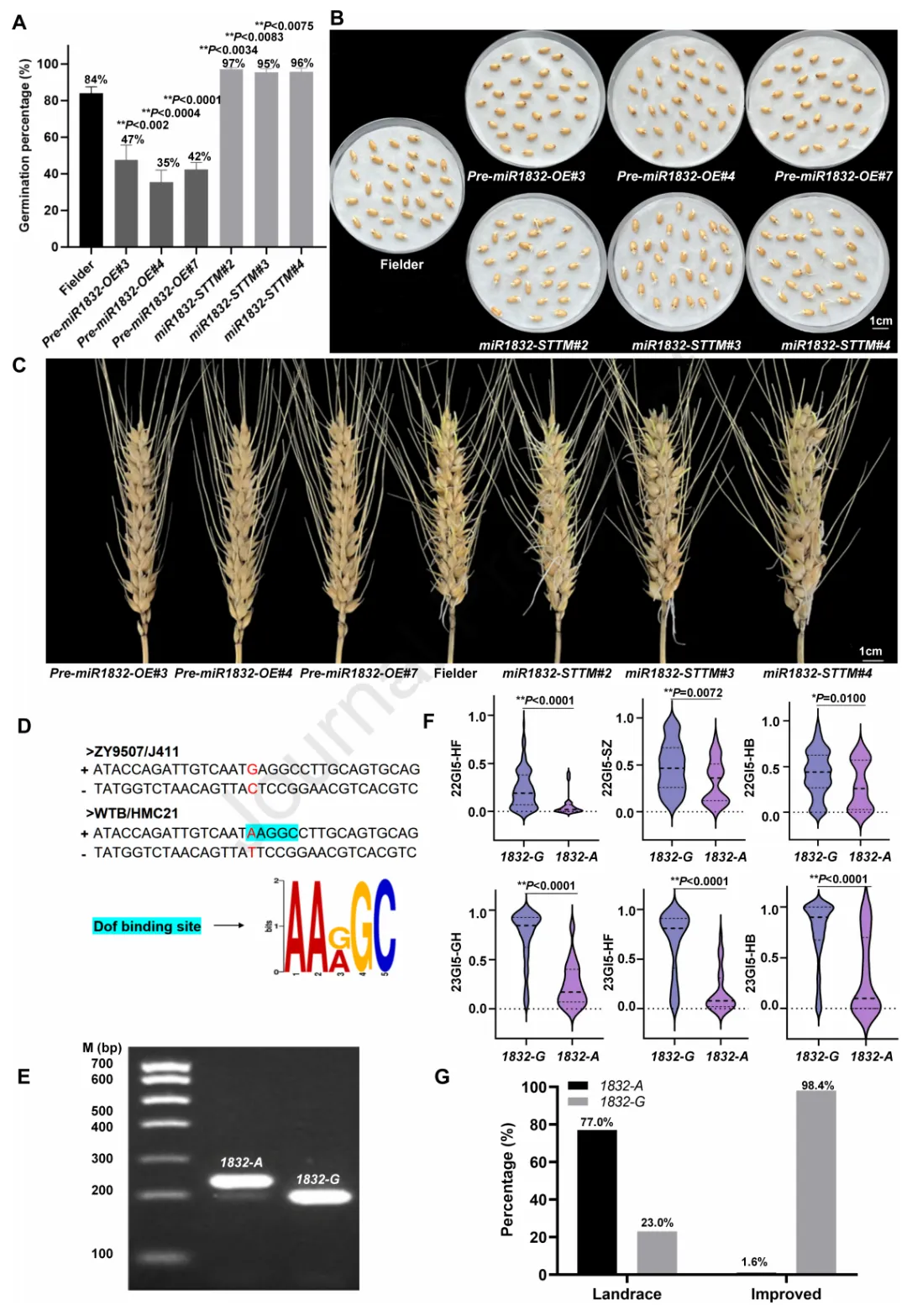
图2.MiR1832正调控小麦种子休眠。(A)“Fielder”、miR1832过表达株系(pre-miR1832-OE)和miR1832沉默株系(miR1832-STTM)小麦种子吸胀3天的发芽率(GP),数值为平均值±标准差(n=3);采用Student’s t检验分析统计显著性(**P<0.01)。(B)“Fielder”、pre-miR1832-OE和miR1832-STTM小麦种子吸胀3天的萌发表型。(C)“Fielder”、pre-miR1832-OE和miR1832-STTM小麦穗吸胀5天的萌发表型。(D)弱休眠品种(“ZY9507”、“J411”)与强休眠品种(“WTB”、“HMC21”)的miR1832序列比对;青色方框代表Dof结合位点,红色碱基代表突变位置。(E)采用2.0%琼脂糖凝胶电泳检测miR1832的酶切扩增多态性序列(CAPS)标记1832。(F)160个小麦品种(160WVs)中携带1832-A和1832-G等位基因的种子在不同环境下(22GI5-HF、22GI5-SZ、22GI5-HB、22GI5-GH、23GI5-HF、23GI5-HB)的发芽指数(GI)差异;1832-A/G:对应miR1832启动子-670bp处A/G核苷酸突变的CAPS标记1832等位基因;采用Student’s t检验分析统计显著性(*P<0.05,**P<0.01)。(G)地方品种和改良品种中miR1832的等位基因频率分布。
05
TaDof-2D直接结合miR1832启动子的A等位基因位点并抑制其转录
强休眠品种“WTB”和“HMC21”的miR1832启动子区-670bp处的A位点被预测为Dof转录因子的结合位点。为筛选能结合该位点的上游转录因子,本研究扩增了包含A和G等位基因的、-670bp位点两侧各45bp的序列(分别命名为miR1832-A和miR1832-G),并将miR1832-A克隆至pAbAi载体构建诱饵系统,进行酵母单杂交文库筛选,共鉴定出35个阳性克隆,其中包含3个Dof转录因子:TaDof-2D(TraesCS2D02G100300)、TaDof-3A(TraesCS3A02G180600)和TaDof-4D(TraesCS4D02G285100)。对常温和低温处理的“WTB”种子进行转录分析,结果显示TaDof-2D的表达受低温显著上调,而TaDof-3A和TaDof-4D的表达无显著变化。
酵母单杂交实验表明,TaDof-2D、TaDof-3A和TaDof-4D均可结合miR1832-A序列,但无法结合miR1832-G序列(图3A、B)。双荧光素酶(LUC)报告基因实验进一步证实了等位基因特异性调控:仅TaDof-2D可显著降低含A等位基因的miR1832启动子的转录活性,而TaDof-2D、TaDof-3A和TaDof-4D均无法调控含G等位基因的miR1832启动子的转录活性(图3C-E)。此外,含A等位基因的miR1832启动子转录活性显著高于含G等位基因的启动子(图3E),这与1832-A基因型小麦品种中miR1832高表达的结果一致。为验证该调控作用的直接性,本研究进行了电泳迁移率变动分析(EMSA),结果显示仅TaDof-2D可特异性结合包含A等位基因的5”-AAGGC-3”基序,而TaDof-2D、TaDof-3A和TaDof-4D均无法结合包含G等位基因的miR1832-G基序(图3F)。综上,TaDof-2D可直接结合miR1832启动子的A等位基因位点,抑制其表达。结合低温处理可上调“WTB”种子中TaDof-2D的表达、下调miR1832表达的结果,本研究推测低温通过上调TaDof-2D的表达,抑制miR1832的转录,进而诱导小麦种子休眠解除。
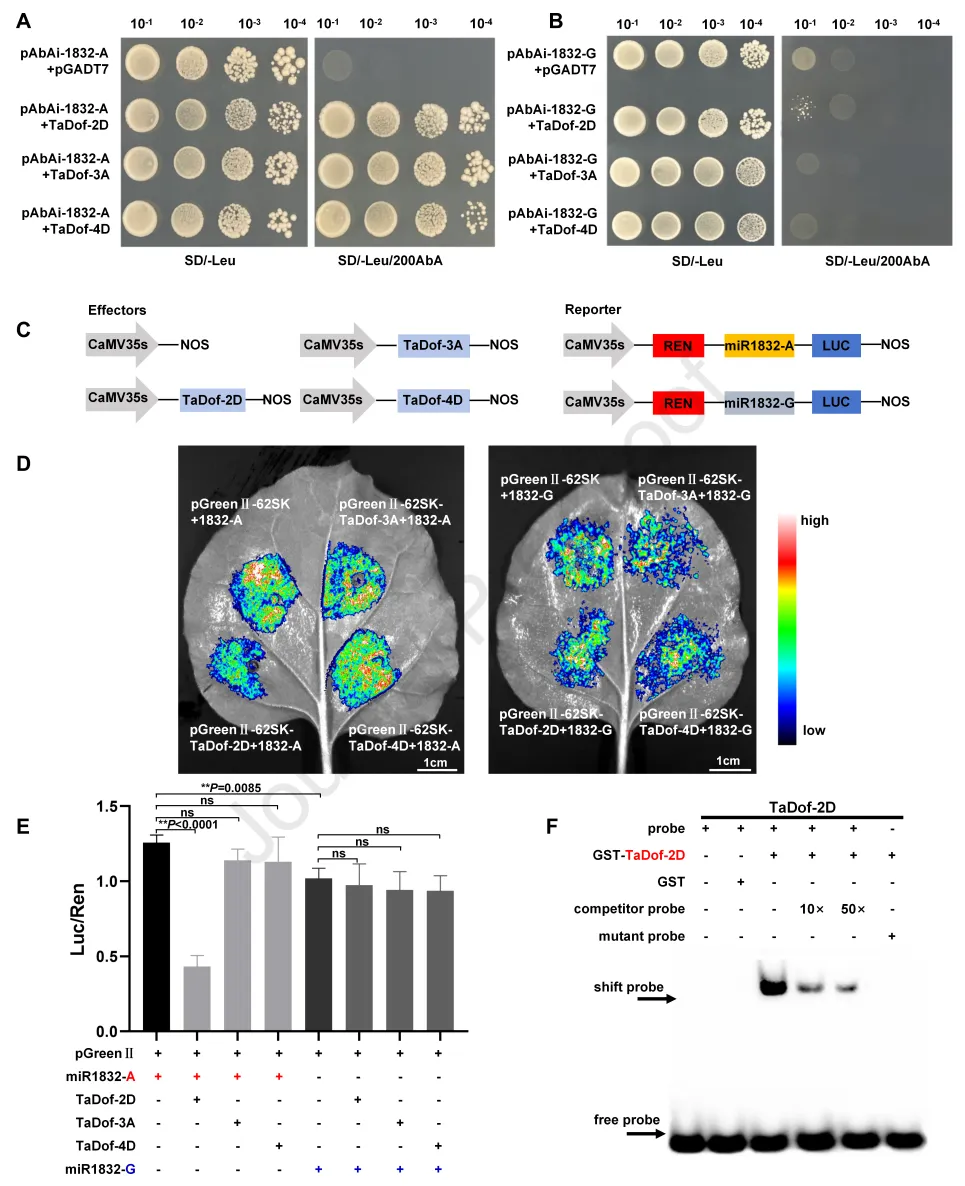
图3. TaDof-2D抑制miR1832转录。(A)酵母单杂交实验(Y1H)显示,TaDof-2D、TaDof-3A和TaDof-4D可结合含A等位基因的miR1832启动子;SD/-Leu:缺亮氨酸合成缺陷培养基;AbA:金担子素A。(B)酵母单杂交实验显示,TaDof-2D、TaDof-3A和TaDof-4D无法结合含G等位基因的miR1832启动子。(C)双荧光素酶(LUC)报告基因实验所用的报告基因和效应元件结构。(D-E)瞬时双荧光素酶报告基因实验显示,TaDof-2D抑制含A等位基因的miR1832启动子转录活性;上图为烟草叶片中的LUC信号,下图为LUC/海肾荧光素酶(REN)比值;数值为平均值±标准差(n=3);采用Student’s t检验分析统计显著性(**P<0.01);ns:无显著差异。(F)电泳迁移率变动分析(EMSA)检测TaDof-2D与miR1832启动子中Dof结合基序AAGGC的结合能力;结合反应中添加10倍和50倍未标记探针作为竞争物;miR1832-A:含A等位基因的miR1832启动子片段;miR1832-G:含G等位基因的miR1832启动子片段;GST:谷胱甘肽S-转移酶。
为验证TaDof-2D对小麦种子休眠的调控作用,本研究在1832-G基因型品种“Fielder”和1832-A基因型品种“WTB”中过表达TaDof-2D的编码序列,分别获得3个“Fielder”背景的过表达株系(Dof-2D-G-OE#2、Dof-2D-G-OE#4、Dof-2D-G-OE#5)和3个“WTB”背景的过表达株系(Dof-2D-A-OE#3、Dof-2D-A-OE#5、Dof-2D-A-OE#6)。萌发实验表明,与野生型“WTB”相比较,Dof-2D-A-OE#3/5/6株系的发芽率显著升高(休眠性降低);而Dof-2D-G-OE#2/4/5株系与野生型“Fielder”的发芽率无显著差异(图4A-F)。此外,本研究在1832-G基因型品种“J411”中获得了TaDof-2D的甲基磺酸乙酯诱变突变体(dof-2d),该突变体在第382位氨基酸处发生G/A单核苷酸突变,导致翻译提前终止;萌发实验显示,dof-2d突变体与野生型“J411”的发芽率无显著差异(图4G-I)。表达检测表明,与野生型“WTB”相比,Dof-2D-A-OE#3/5/6株系中miR1832的表达量显著下调;而Dof-2D-G-OE#2/4/5株系与“Fielder”、dof-2d突变体与“J411”中miR1832的表达量均无显著差异。实时qRT-PCR分析了TaDof-2D和miR1832在“WTB”根、茎、叶、穗和种子中的表达模式,结果显示TaDof-2D在根和茎中表达量较高,在叶、穗和种子中表达量较低;而miR1832的表达模式则与之相反。以上结果证实,TaDof-2D通过特异性结合miR1832启动子的A等位基因位点抑制其转录,进而降低小麦种子的休眠性。
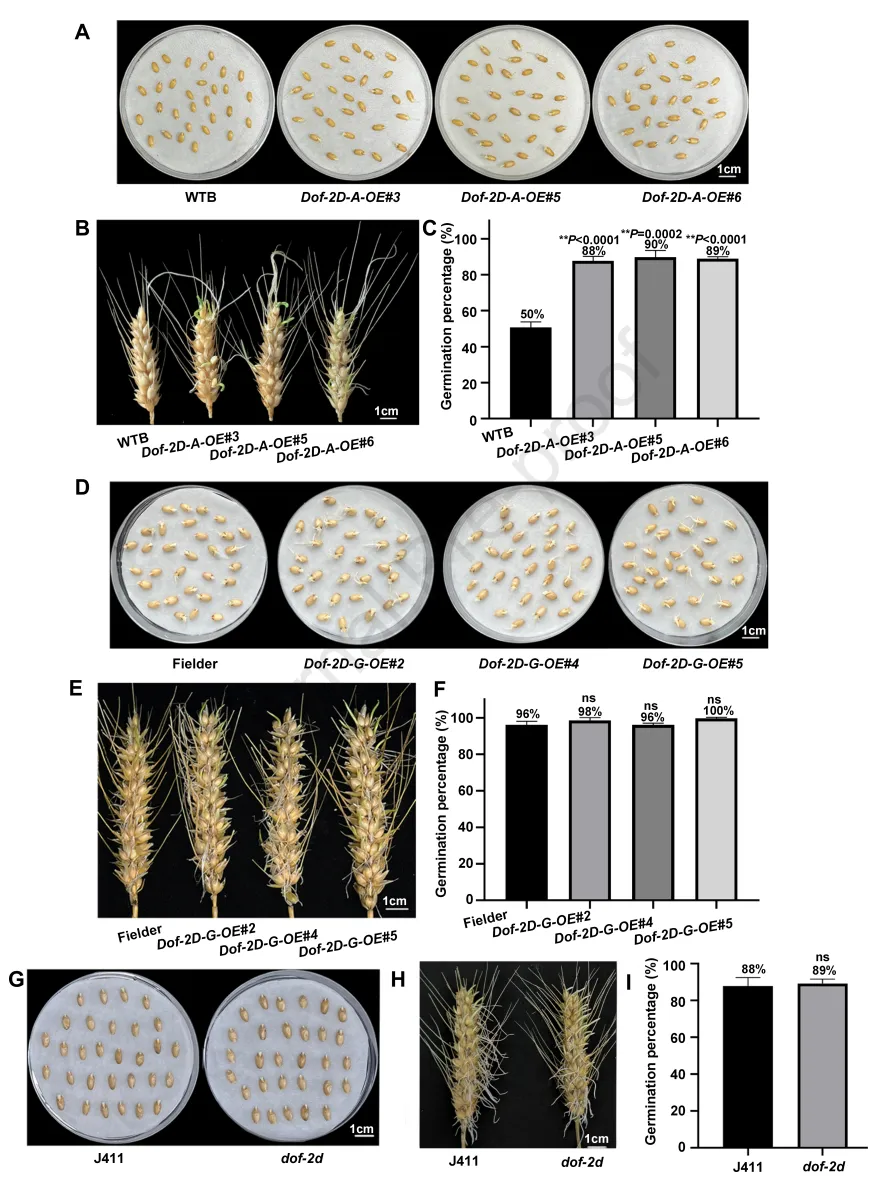
图4. TaDof-2D通过结合miR1832启动子的A位点调控小麦种子休眠。(A)“WTB”和Dof-2D-A-OE#3/5/6种子吸胀3天的萌发表型。(B)“WTB”和Dof-2D-A-OE#3/5/6穗吸胀5天的萌发表型。(C)“WTB”和Dof-2D-A-OE#3/5/6种子的发芽率,数值为平均值±标准差(n=3);采用Student’s t检验分析统计显著性(**P<0.01)。(D)“Fielder”和Dof-2D-G-OE#2/4/5种子吸胀3天的萌发表型。(E)“Fielder”和Dof-2D-G-OE#2/4/5穗吸胀5天的萌发表型。(F)“Fielder”和Dof-2D-G-OE#2/4/5种子的发芽率,数值为平均值±标准差(n=3)。(G)“J411”和dof-2d种子吸胀3天的萌发表型。(H)“J411”和dof-2d穗吸胀5天的萌发表型。(I)“J411”和dof-2d种子的发芽率,数值为平均值±标准差(n=3);ns,无显著差异。
06
TaP450-7A是miR1832的靶标基因
为鉴定miR1832的靶标基因,本研究结合转录组测序结果与psRNATarget靶标预测,筛选出3个潜在靶标基因:乙烯响应转录因子基因TaERF-1D(TraesCS1D02G221500)、NAC转录因子基因TaNAC-4A(TraesCS4A02G219700)和细胞色素P450基因TaP450-7A(TraesCS7A02G455300)。表达分析显示,与miR1832相反,这3个基因在低温处理的“WTB”种子中均显著上调表达。本研究进一步检测了这3个基因在新鲜收获的miR1832过表达株系、沉默株系和野生型“Fielder”种子中的表达模式,结果显示与“Fielder”相比,TaP450-7A在miR1832过表达株系中显著下调表达,在miR1832沉默株系中显著上调表达;而TaERF-1D和TaNAC-4A在过表达、沉默株系与野生型中的表达量均无显著差异。以上结果表明,TaP450-7A可能是miR1832的关键靶标基因。本研究通过5'端cDNA快速扩增技术,定位了miR1832对TaP450-7A mRNA的切割位点(图5A)。为进一步在体内验证miR1832对TaP450-7A的调控作用,本研究对TaP450-7A进行同义突变,获得miR1832抗性版本(mTaP450-7A)(图5B),该突变体可避免被miR1832靶向切割。烟草叶片瞬时表达的双荧光素酶实验表明,miR1832的表达可显著抑制TaP450-7A-LUC报告基因的信号积累,而对mTaP450-7A-LUC报告基因的活性无显著抑制作用(图5C-E)。qRT-PCR检测显示,1832-A基因型小麦品种中TaP450-7A的表达量显著低于1832-G基因型品种。以上结果证实,miR1832可在体内直接靶向切割TaP450-7A的mRNA。
为验证TaP450-7A对小麦种子休眠的调控作用,本研究在“Fielder”中过表达TaP450-7A,获得3个过表达株系(P450-7A-OE#3、P450-7A-OE#6、P450-7A-OE#7)。与“Fielder”相比,3个过表达株系中TaP450-7A的表达量均显著上调;萌发实验表明,过表达株系的发芽率(P450-7A-OE#3为98%、P450-7A-OE#6为97%、P450-7A-OE#7为100%)显著高于“Fielder”(85%)(图5F-H)。此外,本研究在“J411”中获得了TaP450-7A的甲基磺酸乙酯诱变突变体(p450-7a),该突变体在第19位氨基酸处发生G/A单核苷酸突变,导致翻译提前终止;萌发实验显示,p450-7a突变体的发芽率(50%)显著低于野生型“J411”(90%)(图5I-K)。综上,TaP450-7A负调控小麦种子休眠,而miR1832正调控小麦种子休眠,证实了二者间的靶向调控关系。
为探究miR1832与TaP450-7A的遗传调控关系,本研究构建了miR1832与TaP450-7A的过表达杂交株系(pre-miR1832-OE#4/P450-7A-OE#7)。萌发实验表明,杂交株系可逆转miR1832过表达株系的强休眠表型,使种子休眠水平恢复至野生型水平(图5L、M)。此外,本研究检测了不同TaDof-2D转基因小麦品种中TaP450-7A的表达量,结果显示与野生型“WTB”相比,Dof-2D-A-OE#3/5/6株系中TaP450-7A的表达量显著上调;而Dof-2D-G-OE#2/4/5株系与“Fielder”、dof-2d突变体与“J411”中TaP450-7A的表达量均无显著差异。以上结果表明,在Dof-2D-A-OE株系中,TaDof-2D通过特异性结合miR1832启动子的A等位基因位点,抑制miR1832的表达,进而上调TaP450-7A的表达,最终降低小麦种子的休眠。
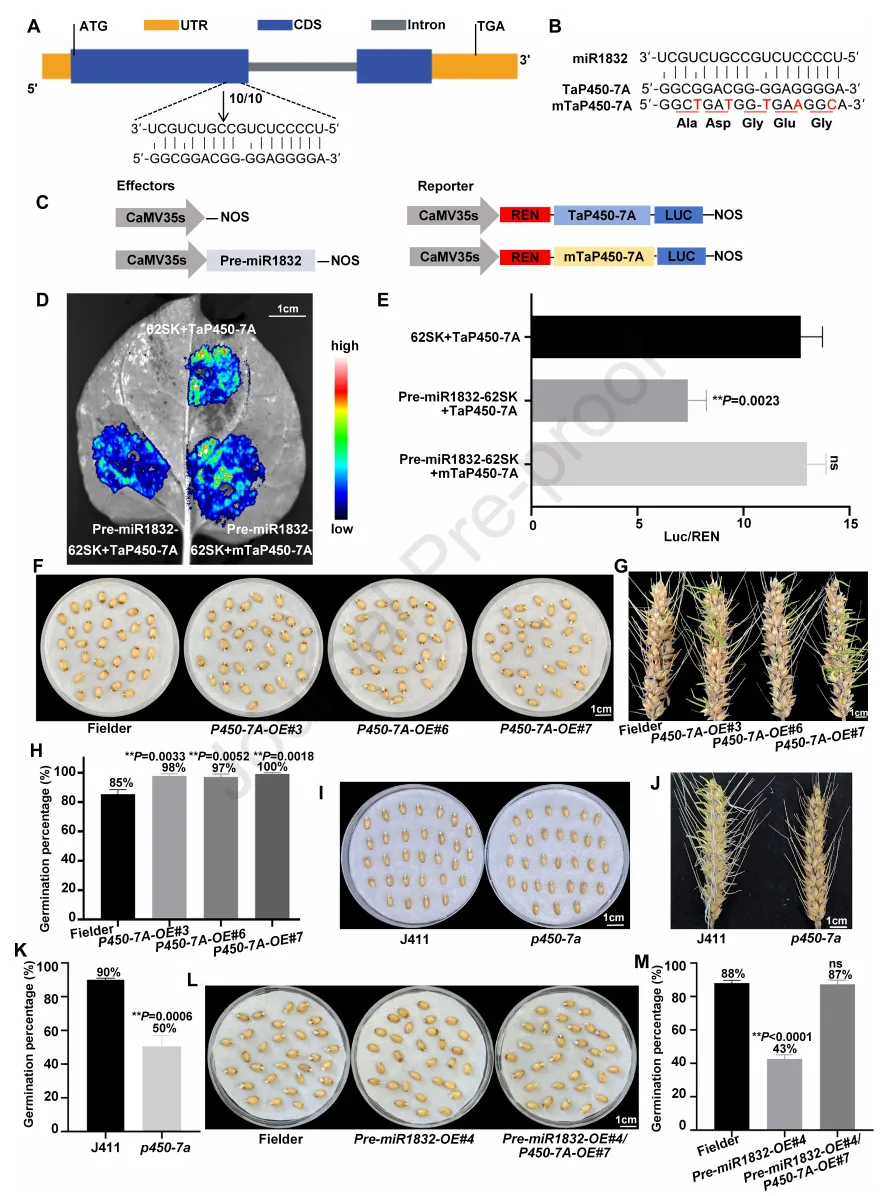
图5. miR1832的靶标验证及功能分析。(A)5′-cDNA快速扩增(5′-RACE)实验显示miR1832介导的TaP450-7A切割位点;黄色、蓝色和灰色方框分别代表非翻译区(UTR)、外显子和内含子。(B)通过同义突变获得miR1832抗性版本(mTaP450-7A)的示意图。(C)双荧光素酶(LUC)报告基因实验所用的报告基因和效应元件结构;CaMV35S,花椰菜花叶病毒35S启动子;Nos,胭脂碱合成酶终止子。(D)转染报告载体和效应/报告载体的本氏烟草组织中的LUC活性。(E)本氏烟草中miR1832与TaP450-7A靶标关系的LUC实验验证,数值为平均值±标准差(n=3)。(F)“Fielder”和P450-7A-OE#3/6/7种子吸胀3天的萌发表型。(G)“Fielder”和P450-7A-OE#3/6/7穗吸胀5天的萌发表型。(H)“Fielder”和P450-7A-OE#3/6/7种子的发芽率,数值为平均值±标准差(n=3)。(I)“J411”和p450-7a种子吸胀3天的萌发表型。(J)“J411”和p450-7a穗吸胀5天的萌发表型。(K)“J411”和p450-7a种子的发芽率,数值为平均值±标准差(n=3)。(L)“Fielder”、pre-miR1832-OE#4和pre-miR1832-OE#4/P450-7A-OE#7种子吸胀3天的萌发表型,数值为平均值±标准差(n=3)(M)“Fielder”、pre-miR1832-OE#4和pre-miR1832-OE#4/P450-7A-OE#7种子的发芽率,数值为平均值±标准差(n=3);采用Student’s t检验分析统计显著性(**P<0.01)。ns,无显著差异。
07
TaDof-2D-miR1832-TaP450-7A调控模块通过调控α-淀粉酶活性及脱落酸、赤霉素通路介导低温诱导的种子休眠解除
小麦种子发育中后期遭遇低温可诱导晚熟α-淀粉酶基因表达,进而在高湿条件下促进种子萌发;且赤霉素和脱落酸可通过拮抗作用调控晚熟α-淀粉酶基因的表达。本研究推测,脱落酸、赤霉素和α-淀粉酶(包括晚熟α-淀粉酶)共同参与低温诱导的“WTB”种子休眠解除。表达分析显示,与常温处理相比,低温处理的“WTB”种子中6个晚熟α-淀粉酶基因(TraesCS6A02G334200,TraesCS6B02G349700,TraesCS6B02G349500,TraesCS6B02G364900,TraesCS6B02G349800,TraesCS6B02G364800)表达显著上调,2个α-淀粉酶抑制基因(TaCM1-7D、TaCM2-7B)表达显著下调;同时,4个赤霉素信号通路基因(TaGID1-1B、TaGID1-1D、TaGASR7-7B、TaGASR7-7D)和2个脱落酸分解代谢基因(TaABA8'OH2-5B、TaABA8'OH2-5D)表达显著上调,2个脱落酸信号通路基因(TaABI5-3B、TaABI5-3D)表达显著下调;实时qRT-PCR验证了这些基因的表达模式。生理指标检测显示,与常温处理相比,低温处理的“WTB”种子中GA1、GA3含量和α-淀粉酶活性显著升高,脱落酸含量显著降低(图1D-F)。综上,小麦种子发育中后期遭遇低温,可通过调控晚熟α-淀粉酶的积累及赤霉素、脱落酸的生物合成、分解代谢与信号通路,诱导种子休眠解除。
为进一步验证TaDof-2D-miR1832-TaP450-7A调控模块是否通过影响赤霉素、脱落酸信号通路关键基因的表达,介导低温诱导的种子休眠解除,本研究测定了吸胀24h的miR1832过表达株系(pre-miR1832-OE#4)、TaP450-7A过表达株系(P450-7A-OE#7)和野生型“Fielder”种子中的赤霉素、脱落酸含量及α-淀粉酶活性。检测到小麦种子中存在两条主要的赤霉素生物合成通路:非13-羟化通路(包括GA15、GA9、GA24、GA4)和早期13-羟化通路(包括GA53、GA44、GA19、GA20、GA1、GA5、GA3)。结果显示,与“Fielder”相比,TaP450-7A过表达株系中早期13-羟化通路的关键前体物质(GA53、GA44、GA19、GA20)和活性产物(GA1、GA3)含量显著升高,而miR1832过表达株系中上述物质含量显著降低(图6A)。此外,TaP450-7A过表达株系中脱落酸含量显著降低、α-淀粉酶活性显著升高,而miR1832过表达株系则呈现相反的趋势(图6B、C)。以上结果为miR1832-TaP450-7A通过调控脱落酸/赤霉素平衡调控小麦种子休眠提供了有力证据。
本研究进一步分析了吸胀24h的pre-miR1832-OE#4、P450-7A-OE#7和“Fielder”种子中脱落酸、赤霉素通路相关基因的表达模式。结果显示,与“Fielder”相比,pre-miR1832-OE#4种子中赤霉素生物合成基因(TaGA20ox1)、赤霉素信号通路基因(TaGASR7-7B、TaGASR7-7D、TaGID1-1B、TaGID1-1D)和脱落酸分解代谢基因(TaABA8'OH-5B、TaABA8'OH-5D)的表达量显著下调,而脱落酸信号通路基因(TaABI5-3B、TaABI5-3D)的表达量显著上调;同时,α-淀粉酶基因TaAmy1-6B的3个同源基因表达量显著下调,α-淀粉酶抑制基因(TaCMI-7D、TaCM2-7B)表达量显著上调。而P450-7A-OE#7种子中上述基因的表达模式与pre-miR1832-OE#4完全相反。综上,TaDof-2D-miR1832-TaP450-7A调控模块通过调控α-淀粉酶活性及赤霉素、脱落酸信号通路,介导低温诱导的小麦种子休眠解除。
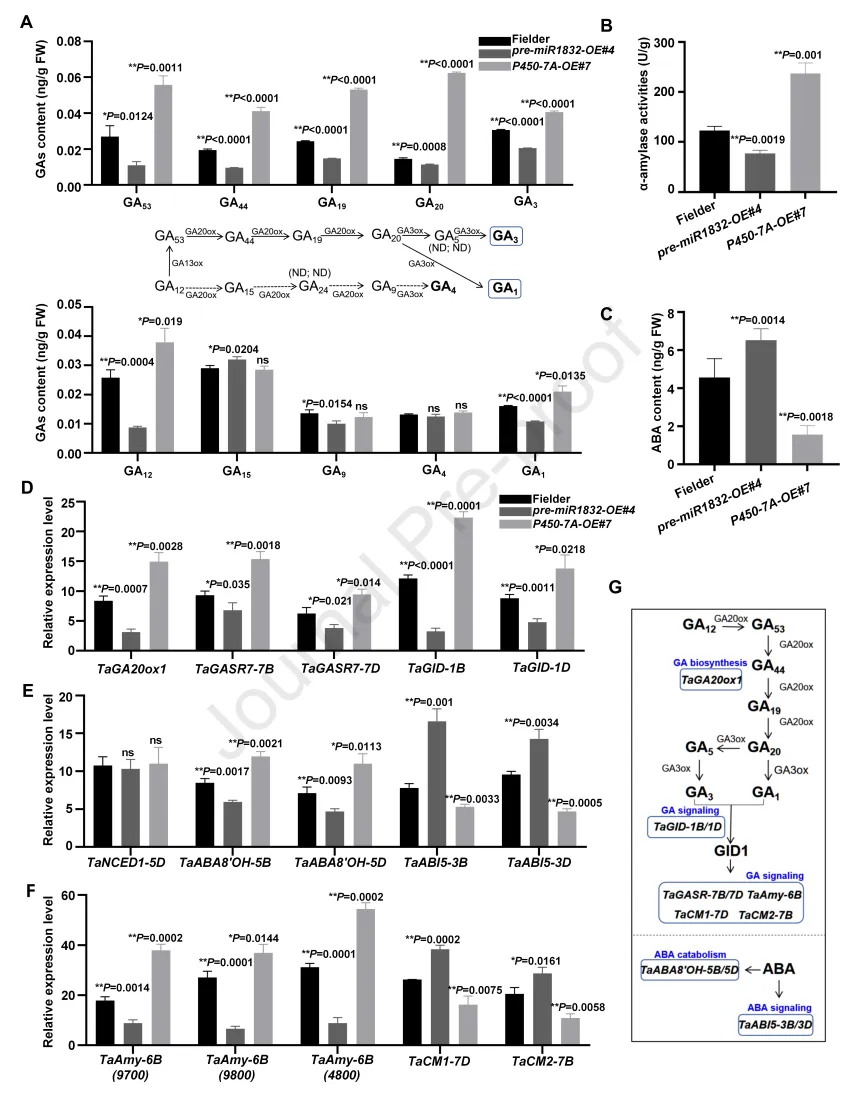
图6. miR1832-TaP450-7A模块通过影响α-淀粉酶活性及脱落酸、赤霉素通路促进低温诱导的种子休眠解除。(A)“Fielder”、pre-miR1832-OE#4和P450-7A-OE#7种子吸胀24h后的赤霉素含量,图示为两条主要的赤霉素生物合成通路:非13-羟化通路(包括GA15[赤霉素A15]、GA9[赤霉素A9]、GA24[赤霉素A24]、GA4[赤霉素A4])和早期13-羟化通路(包括GA53[赤霉素A53]、GA44[赤霉素A44]、GA19[赤霉素A19]、GA20[赤霉素A20]、GA1[赤霉素A1]、GA5[赤霉素A5]、GA3[赤霉素A3]);FW:鲜重;ox:氧化酶;ND:未检测到;数值为平均值±标准差(n=3)。(B-C)“Fielder”、pre-miR1832-OE#4和P450-7A-OE#7种子吸胀24h后的脱落酸含量和α-淀粉酶活性;FW:鲜重;数值为平均值±标准差(n=3)。(D-F)“Fielder”、pre-miR1832-OE#4和P450-7A-OE#7种子吸胀24h后α-淀粉酶生物合成及脱落酸、赤霉素信号通路关键基因的表达水平;基因ID:TraesCS5D02G038800(TaNCED1)、TraesCS5B02G236500(TaABA8'OH-5B)、TraesCS5D02G244900(TaABA8'OH-5D)、TraesCS3B02G404400(TaABI5-3B)、TraesCS3D02G364900(TaABI5-3D)、TraesCS3D02G393900(TaGA20ox1)、TraesCS7B02G115300(TaGASR7-7B)、TraesCS7A02G208100(TaGASR7-7A)、TraesCS1B02G265900(TaGID-1B)、TraesCS1D02G254500(TaGID-1D)、TraesCS6B02G349700(TaAmy1-6B)、TraesCS6B02G349800(TaAmy1-6B)、TraesCS6B02G364800(TaAmy1-6B)、TraesCS7D02G168000(TaCM1-7D)、TraesCS7B02G072000(TaCM2-7B);数值为平均值±标准差(n=3)。(G)赤霉素和脱落酸生物合成、代谢及信号转导通路中关键基因的作用位置示意图。采用Student’s t检验分析统计显著性(*P<0.05,**P<0.01);ns,无显著差异。
结论




END


